
7 Chapter 7: Modern Human Origins and Agriculture
Amy Rector
By the end of this chapter, you should be able to:
- Identify the skeletal and behavioral traits that represent modern Homo sapiens.
- Critically evaluate different types of evidence for the origin of our species in Africa, and our expansion around the world.
- Understand how the human lifestyle changed when people transitioned from foraging to agriculture.
Defining Characteristics of Archaic Homo Sapiens
Archaic Homo sapiens share our species name but are distinguished by the term “archaic” as a way of recognizing both the long period of time between their appearance and ours, as well as the way in which human traits have continued to evolve over time—making archaic Homo sapiens look slightly different from us today, despite technically being considered the same species. Living throughout Europe and Asia during the Middle Pleistocene, archaic Homo sapiens are considered, in many ways, transitional between Homo erectus and modern Homo sapiens (see chart). All archaic Homo sapiens share the defining trait of an increased brain size—specifically a brain of at least 1,100 cc and averaging 1,200 cc—but are also characterized by significant regional and temporal variations. Because of these variations, scientists disagree on whether these fossils represent a single, variable species or multiple, closely related species (sometimes called Homo antecessor, Homo heidelbergensis, Homo georgicus, Homo neanderthalensis, and Homo rhodesiensis). For simplicity, we are going to lump them all together under the heading of archaic Homo sapiens and discuss them as a unit, with the exception of a particularly unique and well-known population living in Europe and West Asia known as the Neanderthals, which we will examine separately.
|
Trait |
Homo erectus |
Archaic Homo sapiens |
Anatomically Modern Homo sapiens |
|
Average Brain Size |
900 cc |
1,200 cc (1,500 cc when including Neanderthals) |
1,400 cc |
|
Skull Shape |
Long and low Angular |
Intermediate |
Short and high |
|
Forehead |
Absent |
Emerging |
Present |
|
Nasal Region |
Projecting nasal bones (bridge of the nose), no midfacial prognathism |
Wider nasal aperture and midfacial prognathism |
Narrower nasal aperture, no midfacial prognathism |
|
Chin |
Absent |
Absent |
Present |
|
Other Facial Features |
Large brow ridge and large projecting face |
Intermediate |
Small brow ridge and retracted face |
|
Other Skull Features |
Nuchal torus, sagittal keel, thick cranial bone |
Projecting occipital bone, often called occipital bun in Neanderthals; intermediate thickness of cranial bone |
Small bump on rear of skull, if anything; thin cranial bone |
|
Dentition |
Large teeth, especially front teeth |
Slightly smaller teeth; front teeth still large; retromolar gap in Neanderthals |
Smaller teeth |
|
Postcranial Features |
Robust bones of skeleton |
Robust bones of skeleton |
More gracile bones of skeleton |
When comparing Homo erectus, archaic Homo sapiens, and anatomically modern Homo sapiens across several anatomical features, one can see quite clearly that archaic Homo sapiens are intermediate in their physical form. To identify a set of features that is unique to archaic Homo sapiens is a challenging task, due to both individual variation—these developments were not all present to the same degree in all individuals—and the transitional nature of their features. Neanderthals are an exception, as they have several clearly unique traits that make them notably different from modern Homo sapiens as well as their closely related archaic cousins.
Due to the transitional nature of archaic Homo sapiens, identifying the time period they are associated with is complex. Generally, it is agreed upon that archaic Homo sapiens lived between 600,000 and 200,000 years ago. But regionally this varies, with considerable overlap between Homo erectus on the early end of the spectrum and modern Homo sapiens and Neanderthals on the latter end. The earliest-known archaic Homo sapiens fossils tentatively date to about 600,000 years ago in Africa, whereas archaic Homo sapiens fossils in Asia appear around 300,000 years ago and in Europe around 350,000 years ago (and potentially as early as 600,000 years ago). The end point of archaic Homo sapiens is also problematic since it largely depends upon when the next subspecies of Homo sapiens appears and the classification of highly intermediate specimens. For example, in Africa, the end of archaic Homo sapiens is met with the appearance of modern Homo sapiens, while in Europe it is the appearance of Neanderthals that is traditionally seen as the end of archaic Homo sapiens.
Archaic Homo sapiens mark an important chapter in the human lineage, bridging more ancestral forms, such as Homo erectus, with modern Homo sapiens. During this period of climatic transition and fluctuation, archaic Homo sapiens mirror the challenges of their environments. Showing increasing regional variation due to the need for local adaptation, there is no single archetype for this group but, rather, multiple variations; their transitional nature is one of their key defining characteristics.
Neanderthals
One particularly well-known population of archaic Homo sapiens are the Neanderthals, named after the site where they were first discovered in the Neander Valley, or “thal” in German, located near Dusseldorf, Germany. Popularly known as the stereotypical “cavemen,” recent research is upending long-held beliefs about this group. Neanderthals have long been thought to be dumb brutes who were, behaviorally speaking, not much different from apes. However, today, scientists agree that Neanderthal behavior was increasingly complex and nuanced, far beyond what was exhibited by even other archaic Homo sapiens.
It is important to understand why Neanderthals are typically separated from other archaic Homo sapiens. Unlike the rest of archaic Homo sapiens, Neanderthals are easily defined and identified in many ways. There is a clear geographic boundary of where Neanderthals lived: western Europe, the Middle East, and western Asia. The time period for when Neanderthals lived is widely accepted as between 150,000 and 35,000 years ago. Additionally, Neanderthals have a unique and distinct cluster of physical characteristics. While a few aspects of Neanderthals are less clear cut and are shared among some archaic Homo sapiens, such as the types of tools they created and used, most attributes of Neanderthals, both anatomically and behaviorally, are unique to them.
Neanderthals lived during some of the coldest times during the last Ice Age and at far northern latitudes. This means Neanderthals were living very close to the glacial edge, and not in a more temperate region of the globe, like many of their archaic Homo sapiens relatives. Their range likely expanded and contracted along with European glacial events, moving into the Middle East during glacial events when Europe became even cooler, and when the animals they hunted would have moved for the same reason. During interglacials, when Europe warmed a bit, Neanderthals and their prey would have been able to move back into Western Europe.
Many of the Neanderthals’ defining physical features are more extreme and robust versions of traits seen in other archaic Homo sapiens, clustered in this single population (Figure 1). Brain size is one of the Neanderthal features that continues to follow the same patterns as seen with other archaic Homo sapiens, namely an enlargement of the cranial capacity. The average Neanderthal brain size is around 1,500 cc, and the range for Neanderthal brains can extend to upwards of 1,700 cc. The majority of the increase in the brain occurs in the occipital region, or the back part of the brain, resulting in a skull that has a large cranial capacity with a distinctly long and low shape that is slightly wider than previous forms at far back of the skull. Modern humans have a brain size comparable to that of Neanderthals; however, our brain expansion occurred in the frontal region of the brain, not the back, as in Neanderthal brains. This difference is also the main reason why Neanderthals lack the vertical forehead that modern humans possess. They simply did not need an enlarged forehead, because their brain expansion occurred in the rear of their brain. Due to cranial expansion, the back of the Neanderthal skull is less angular (as compared to Homo erectus) and is more rounded, a feature similar to that of modern Homo sapiens.
Another feature that continues the trend noted in previous hominins is the enlargement of the nasal region, or the nose. Neanderthal noses are large and have a wide nasal aperture, which is the opening for the nose. The teeth of the Neanderthals follow a similar pattern seen in the archaic Homo sapiens, which is an overall reduction in size, especially as compared to the extremely large teeth seen in the robust australopithecines. However, while the teeth have continued to reduce, the jaw size does not keep pace, leaving Neanderthals with an interesting situation. Their jaw is oversized for their teeth, leaving a gap between their final molar and the end of their jaw.
Continuing the archaic Homo sapiens trend, Neanderthal brow ridges are prominent but somewhat smaller in size than those of Homo erectus and earlier archaic Homo sapiens. In Neanderthals, the brow ridges are also often slightly less arched than those of other archaic Homo sapiens.
The Neanderthal postcrania are also unique in that they demonstrate increased robusticity in terms of the thickness of bones and body proportions that show a barrel-shaped chest and short, stocky limbs, as well as increased musculature. These body portions are seen across the spectrum of Neanderthals—in men, women, and children.
Many of the unique traits that Neanderthals possess can be attributed to adaptation to the extreme cold environments in which they often lived. For example, Bergmann’s and Allen’s Rules dictate that an increased body mass and short, stocky limbs are common in animals that live in cold conditions. Neanderthals match the predictions of Bergmann’s and Allen’s Rules perfectly. In addition, the Neanderthal skull also exhibits adaptations to the cold. Neanderthals’ large infraorbital foramina allow for larger blood vessels, increasing the volume of blood that is found closest to the skin, which helps to keep the skin warmer. Their enlarged noses may also have been beneficial to have in cold weather due to longer nasal passages and mucus membranes for cold air to travel through before reaching the lungs. It is very uncomfortable and challenging to breathe and exert oneself in exceptionally cold, dry air. The more time the air spends in the mucus membranes, the warmer and more moist the air will be before it reaches the lungs. The Neanderthals’ larger nose has long been thought to have acted as a humidifier, easing physical exertion in their climate, although research on this particular trait continues to be studied and debated.
|
Distinct Neanderthal Anatomical Features |
|
|
Brain Size |
1,500 cc average |
|
Skull Shape |
Long and low |
|
Brow Ridge Size |
Large |
|
Nose Size |
Large, with midfacial prognathism |
|
Dentition |
Reduced, but large jaw size, creating retromolar gap |
|
Occipital Region |
Enlarged occipital region, occipital bun |
|
Other Unique Cranial Features |
Large infraorbital foramina |
|
Postcranial Features |
Short and stocky body, increased musculature, barrel-shaped chest |
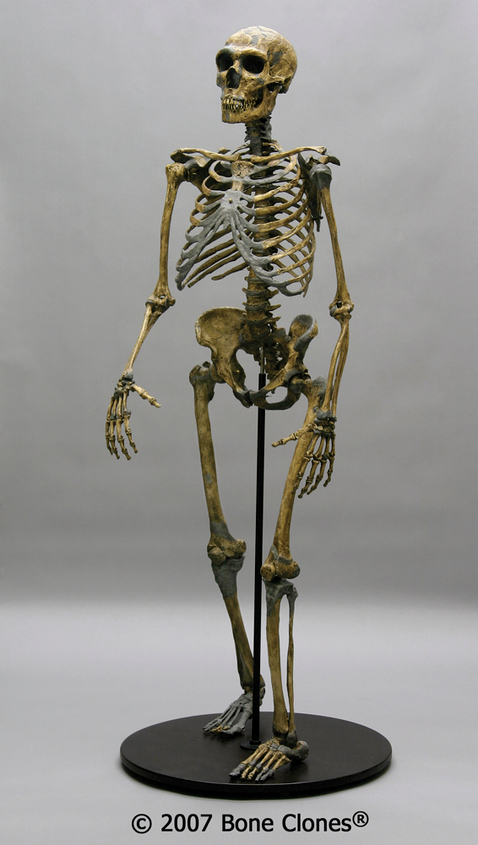
In summary, Neanderthal characteristics are a distinct cluster of features, some of which were apparent in previous hominins and others that were unique. Additionally, it is clear that Neanderthals were specially adapted to a particular environment—a very cold one. A classic example of a Neanderthal with all of the characteristics mentioned above is the La Ferrassie 1 Neanderthal, from France. The skeleton is near complete, which is not necessarily unique among Neanderthal fossils as many partially complete remains have been found, but it does provide us with a lot of information. The La Ferrassie 1 Neanderthal, who was male, had a brain size of around 1640cc and had an extremely large nose and infraorbital foramina. Additionally, the brow ridges are marked in size, and the overall skeleton is robust (Figure 11.7).
What are the benefits or the potential challenges Neanderthals could have faced for being highly specialized to one particular environment, when we know their environment and climate were in flux?
Neanderthal Culture and Lifeways
One key Neanderthal adaptation was their cultural innovations. Cultural innovation is a key way that hominins adapt to their environment. As you recall, the culture of Homo erectus was marked by the development of a bifacial tool, the Acheulean handaxe, which allowed them differential access to meat on animal carcasses when compared to their predecessors. For Homo erectus, the Acheulean handaxe allowed more efficient removal of meat and calculated scavenging. The increase in their body and brain size, along with their more effective tools, allowed them to track predators and snatch their kills sometimes even before the predators themselves had even fed.
Neanderthal tools mark a significant innovation both in tool-making technique and their use. Known as Mousterian tools, after the Le Moustier site in southwest France, the Neanderthal’s toolkit was truly that—a set of tools with specific applications and unique forms for each desired purpose. Mousterian tools were significantly smaller, thinner, and lighter than Acheulean handaxes. The materials used for Mousterian tools were of higher quality, which allowed for both more precise toolmaking and tool reworking when the tools broke or dulled after frequent reuse. The use of higher-quality materials is also indicative of required forethought and planning to acquire them for tool manufacture. It is noteworthy that the Neanderthals, unlike Homo erectus, saved and reused their tools, rather than making new ones each time a tool was needed.
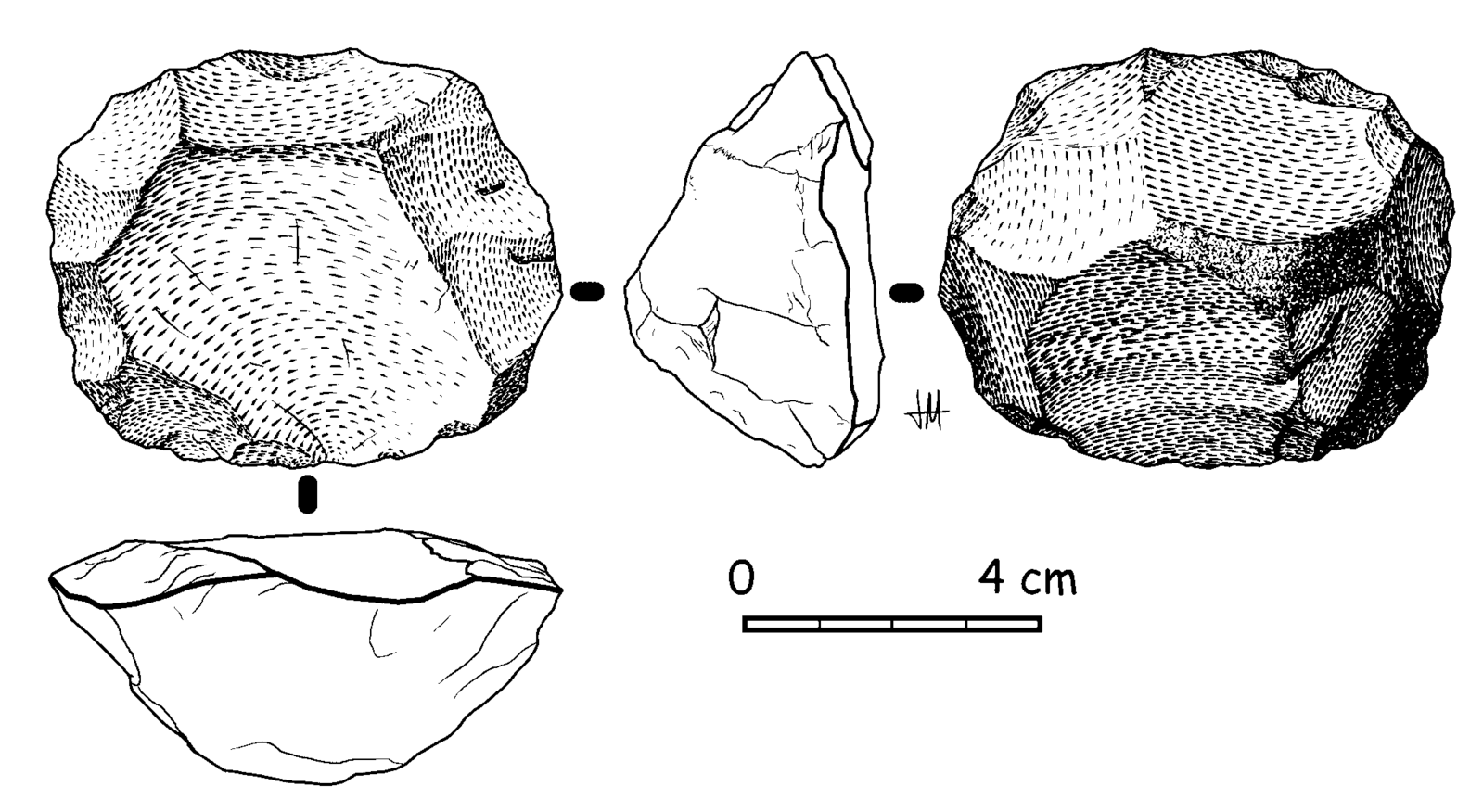
Neanderthal tools were used for a variety of purposes. They would have constructed a tool for each specific task they needed to complete, such as cutting, butchering, woodworking or antler working, and hide working. Additionally, because the Mousterian tools were lighter than previous stone tools, Neanderthals could attach the tool onto a handle, as the stone would not have been too heavy. Neanderthals attached stone blades onto short wood or antler handles to make knives or other small weapons, as well as attached larger blades onto longer shafts to make spears. New research examining tar-covered stones and black lumps at several Neanderthal sites in Europe suggests that Neanderthals may have been making tar by distilling it from birch tree bark, which could have been used to glue the stone tool onto its handle. If Neanderthals were, in fact, manufacturing tar to act as glue, this would predate evidence of modern humans in Africa making tree resin or similar adhesives by nearly 100,000 years! While research on specific applications continues, it is clear that Neanderthal tool manufacturing was much more complex than previous tool-making efforts, requiring skill and patience to carry out.
With their more sophisticated suite of tools, Neanderthals were better armed for hunting than previous hominins and had very robust bodies with larger muscles. The animal remains in Neanderthal sites show that unlike earlier archaic Homo sapiens, Neanderthals were very effective hunters who were able to kill their own prey. Oftentimes, this included very large animals like deer, horses, and bovids (relatives of the cow). In fact, isotopes from Neanderthal bones show that meat was a primary and significant component of their diet, similar to that seen in carnivores like wolves (Bocherens et al. 1999).
Though more sophisticated than the tools of earlier hominins, the Neanderthal spear was not the kind of weapon that would have been thrown; rather, it would have been used in a jabbing fashion (Kortlandt 2002). This may have required Neanderthals to hunt in groups rather than individually, and it almost certainly meant that they would have had to approach their prey quite closely. Remember, the animals living with Neanderthals were also large-bodied due to their adaptations to cold weather. But in addition to large bovids, prey included ibex, seals, rabbits, and pigeons. Though red meat was a critical component of the Neanderthal diet, evidence shows that at times they also ate limpets, mussels, and pine nuts. Tartar examined from Neanderthal teeth in Iraq and Belgium reveal that plant material including wheat, barley, date palms, and tubers were also eaten by Neanderthals and were cooked to make them palatable.
While the close-range style of hunting used by Neanderthals was effective, it also had some major consequences. Many Neanderthal skeletons have been found with significant injuries, which could have caused paralysis or severely limited their mobility. Many of the injuries are to the head, neck, or upper-body. Thomas Berger and Erik Trinkaus (1995) conducted a statistical comparative analysis of Neanderthal injuries compared to those recorded in modern day workers’ compensation reports and found that the closest match was between Neanderthal injuries and those of rodeo workers. Rodeo professionals have a high rate of head and neck injuries that are similar to the Neanderthals’ injuries. What do Neanderthals and rodeo workers have in common? They were both getting very close to large, strong animals, and at times their encounters might have gone awry.
The extensive injuries sustained by Neanderthals are evident in many fossil remains. Shanidar 1 (Figure 3), an adult male found at the Shanidar site in northern Iraq and dating to 45,000 ya, has a lifetime of injuries recorded in his bones. Shanidar 1 sustained—and healed from—an injury to his face that would have likely caused blindness. His lower right arm was missing and his right humerus shows severe atrophy, likely due to disuse. This pattern has been interpreted to indicate a substantial injury that required or otherwise resulted in amputation or wasting away of the lower arm. Additionally, Shanidar 1 suffered from severe arthritis in his feet and bony growths in his inner ear that would have significantly impaired his hearing. He also exhibited extensive anterior tooth wear, matching the pattern of wear found among modern populations who use their teeth as a tool. Rather than an anomaly, the type of injuries evident in Shanidar 1 are similar to those found in many other Neanderthal fossils, revealing injuries likely sustained from hunting large mammals as well as demonstrating a long life of physical activity.
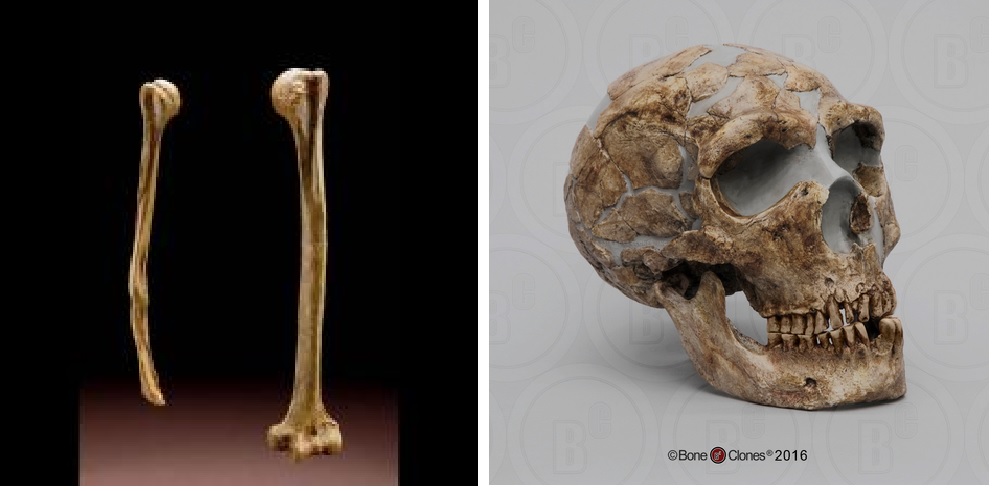
The pattern of injuries is as significant as the fact that Shanidar 1 and other injured Neanderthals often show evidence of having survived their severe injuries. The care expressed in taking care of the sick or injured may have been expressed upon death as well. Full Neanderthal skeletons are not uncommon in the fossil record, and many of these skeletons were so well preserved due to having been placed in deliberate burials. These burials appear intentional, as the graves are dug down a bit, and the bodies found in the graves are in specific positions quite distinct from the natural position the body automatically goes into after death during rigor mortis. Some tantalizing evidence for symbolism and ritual is the frequent occurrence of natural pigments, such as ochre (red) and manganese dioxide (black) in Neanderthal sites. Such pigments could have been used for art, like some of the spectacular cave paintings produced by modern humans who lived in this area after the Neanderthals. However, how these pigments were actually used by Neanderthals themselves is unclear, as there is very little evidence of art or paintings in Mousterian sites. One exception may be the recent discovery in Spain of a perforated shell that appears to be painted with an orange pigment, which may be some of the best evidence of Neanderthal art and jewelry. However, many pigments also have properties that make them good emulsifiers in adhesive (like for attaching a stone tool to a wooden handle) or useful in tanning hides.
The more recent time period in which Neanderthals lived and extensive excavations completed across Europe allow for a much more complete archaeological record from this time period. Additionally, the increased cultural complexity such as complex tools and ritual behaviors expressed by Neanderthals left a more detailed record than previous hominins. Intentional burials enhanced preservation of the dead and potentially associated ritual behaviors. Such evidence allows for a more complete and nuanced picture of this species.

Additional analyses are possible on many Neanderthal finds, due to increased preservation of bone, the amount of specimens that have been uncovered, and the recency in which Neanderthals lived. These additional studies include the examination of dental calculus and even DNA analysis. While limited, samples of Neanderthal DNA have been successfully extracted and analyzed. Studies thus far have identified specific genetic markers that show some Neanderthals were light-skinned and probably red-haired with light eyes. Genetic analyses, different than the typical hominin reconstruction done with earlier species, allow scientists to further investigate soft tissue markers of Neanderthals and other more recent hominin species. These studies and Neanderthal cultural behavior have given scientists a wealth of information to study and offer striking conclusions regarding Neanderthal traits, their physical appearance, and their culture, as reflected in these artists’ reconstructions (Figure 4).
The Neanderthals’ more complicated behavior likely stems, in part, from their larger brains. Evidence shows that raw materials used by Neanderthals came from distances as far away as 100 km. This could indicate a variety of things regarding Neanderthal behavior, including a limited trade network with other Neanderthal groups, or simply a large area scoured by Neanderthals when collecting raw materials. Additionally, we know that Neanderthals lived in groups and may have relied on their group members for survival. Shanidar 1 and the “Old Man at La Chapelle” would have struggled to acquire and consume food on their own, strongly suggesting that they may have been assisted by relatives of other group members. In other nonhuman primates (like chimpanzees) and earlier hominins, injured individuals would have been left on their own, to either survive or perish.
Where Did They Go? The End of Nanderthals
While hominins living in Africa were increasingly successful and ultimately transitioned into modern Homo sapiens, Neanderthals disappear from the fossil record by around 35,000 years ago. The main question that lingers, however, is what happened to them. We know, based on genetics, that modern humans come largely from the modern people who occupied Africa around 300,000 to 100,000 years ago, at the same time that Neanderthals were living in the icy portions of northern Europe and Asia. Modern humans expanded out of Africa by at least 60,000 to 40,000 years ago, rapidly entering areas of Europe and Asia inhabited by Neanderthals and other populations of archaic hominins. Despite intense interest and speculation in fictional works about possible interactions between these two groups, there is very little direct evidence of either peaceful coexistence or aggressive encounters. It is clear, though, that these two closely related hominins shared Europe for thousands of years, and recent DNA evidence suggests that they at least occasionally interbred. Geneticists have found traces of Neanderthal DNA (1% to 4%) in modern humans of European and Asian descent that is not present in modern humans from Africa. This is generally interpreted as indicating limited regional interbreeding with Neanderthals. Interestingly, while some studies suggest interbreeding was often problematic for their offspring, gene flow from Neanderthals may have benefited modern Homo sapiens.
While some interbreeding likely occurred, as a whole, Neanderthals did not survive. What is the cause for their extinction? This question has fascinated many researchers and several possibilities have been suggested that deserve some exploration. Possibilities include the following:
- The climate began changing considerably from the height of Neanderthal expansion. At the time that Neanderthals were disappearing from the fossil record, the climate went through both cooling and warming periods—each of which posed challenges for Neanderthal survival (Defleur and Desclaux 2019; Staubwasser et al. 2018). It has been argued that as temperatures warmed, large-bodied animals, well adapted to cold weather, moved farther north to find colder environments or face extinction themselves. A shifting resource base could have been problematic for continued Neanderthal existence, especially as additional humans, in the form of modern Homo sapiens, began to appear in Europe and were competing with them for available resources.
- It has been suggested that the eruption of a European volcano 40,000 years ago could have put a strain on available plant resources (Golovanova et al. 2010). The eruption would have greatly affected local microclimates, reducing the overall temperature enough to alter the growing season.
- Possible differences in cognitive development may have limited Neanderthals in terms of their creative problem solving. It has been argued that as much as they were biologically specialized for their environment, the nature of their intelligence might not have offered them the creative problem solving skills to rethink their cultural adaptations and innovate ways to adapt their culture when faced with a changing environment (Pearce, Stringer, and Dunbar 2013).
- There is evidence that suggests reproduction may have posed challenges for Neanderthals. Childbirth was thought to have been at least as difficult for female Neanderthals as anatomically modern Homo sapiens (Weaver and Hublin 2009). Female Neanderthals may have become sexually mature at an older age, even older than modern humans. This delayed maturation could have kept the Neanderthal population size small.
- The end of Neanderthal existence also overlaps with modern human expansion into northern Europe and Asia. There is no conclusive direct evidence to indicate that Neanderthals and modern humans lived peacefully side by side, nor that they engaged in brutal warfare, but by studying modern societies, modern humans may not have warmly embraced their close but slightly odd-looking cousins when they first encountered them. It is also completely possible that modern humans gave Neanderthals diseases to which they previously had little to no exposure, causing a mass population decline similar to what happened when the invading Spanish wiped out the Mayan and Aztec populations (Houldcroft and Underdown 2016). Estimates of energy expenditures suggest Neanderthals had slightly higher caloric needs than modern humans. When competing for similar resources, the slightly greater efficiency of modern humans might have helped them experience greater success in the face of competition—at a cost to Neanderthals.
- Finally, less dramatically yet still significantly, even a small but continuous decrease in fertility would have been enough to result in the extinction of Neanderthals (Degioanni et al. 2019).
As Neanderthal populations were fairly small to begin with (estimated between 5,000 and 70,000 individuals), one or a combination of these factors could have easily led to their demise. As more research is conducted, we will likely get a better picture of exactly what led to Neanderthal extinction.
How Do These Fit In? Homo Naledi and Homo Floresiensis
Relatively recently, fossils have been unearthed that have significantly challenged our understanding of the hominin lineage. The fossils of Homo naledi and Homo floresiensis are significant for several reasons, but are mostly known for how they don’t fit the previously held patterns of hominin evolution.
Homo naledi
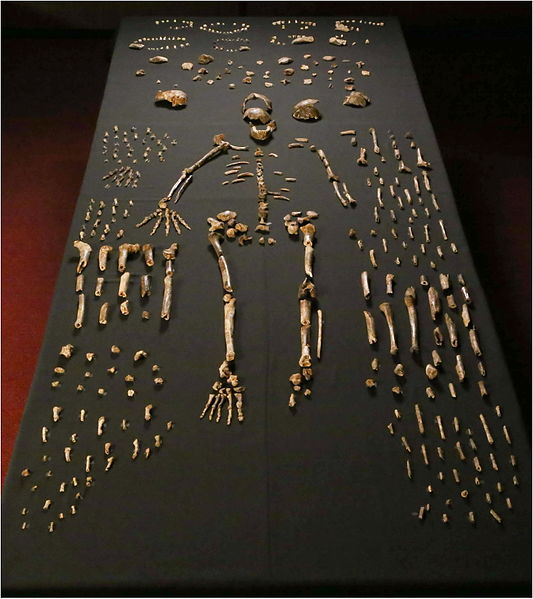
Found in 2013 by recreational spelunkers, a collection of bones was uncovered in a deep cave network in Johannesburg, South Africa. The cave system, known as Rising Star, had been well documented by other cavers; however, it appears few people had ever gone as far into the cave as these spelunkers. The features of Homo naledi are well-documented due to the fairly large sample, which represents individuals of all sexes and a wide range of ages. The skull shape and features are very much like other members of the genus Homo, including features such as a sagittal keel and large brow, like Homo erectus, and a well-developed frontal lobe, similar to modern humans, yet the brain size is significantly smaller than its counterparts, at approximately 500 cc (560 cc for males and 465 cc for females). The teeth also exhibit features of later members of the genus Homo, such as Neanderthals, including a reduction in overall tooth size. Homo naledi also had unique shoulder anatomy and curved fingers, indicating similarities to tree-dwelling primates, which is very different from any other hominin yet found. The date on the species, which was obtained some time after the fossils were first analyzed, indicates that Homo naledi lived between 335,000 and 236,000 years ago. This has been perhaps the greatest shock of all about Homo naledi, as this places this fossil as a contemporary to modern Homo sapiens, despite the very primitive features it retains.
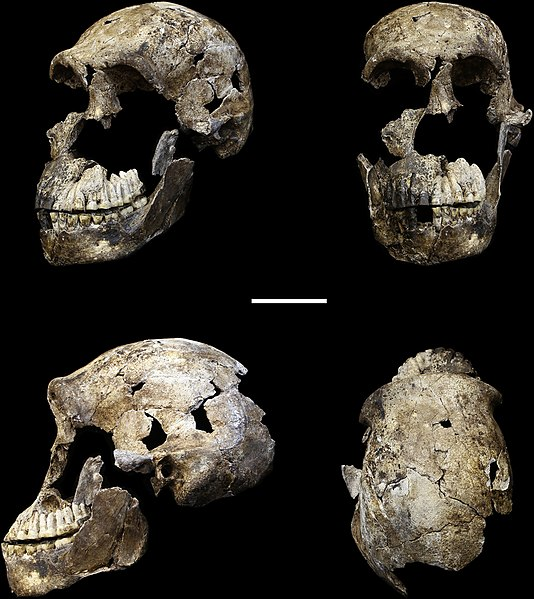
Other remarkable aspects of the find is the placement of the Homo naledi fossils and what it may suggest. To access the site, approximately 80 m from any known cave entrance or opening, a treacherous route would have had to have been taken—including moving through a portion that is just 25 cm wide at some points, known as “Superman’s Crawl.” The only way to get through this section is by crawling on your stomach with one arm by your side and the other raised above your head. Past Superman’s Crawl, a jagged wall known as the Dragon’s Back would have been very difficult to traverse. Below that, a narrow vertical chute would have eventually led down to the area where the fossils were discovered. While geology changes over time and the cave system likely has undergone its fair share, it is not likely that these difficult features arose after Homo naledi lived. This has made scientists curious as to how the bones ended up in the bottom of the cave system in the first place. Some have suggested that Homo naledi deposited the bones there, one way or another. Another competing idea is that a few individuals may have entered the cave system to escape a predator and then got stuck. To account for the sheer number of fossils, this would have had to happen multiple times. In contrast, if Homo naledi did deposit the bones, either through random disposal or intentional burial, this calls into question their symbolic behavior and other cultural traits, including the use of fire, to access a very dark cave system.
Homo floresiensis

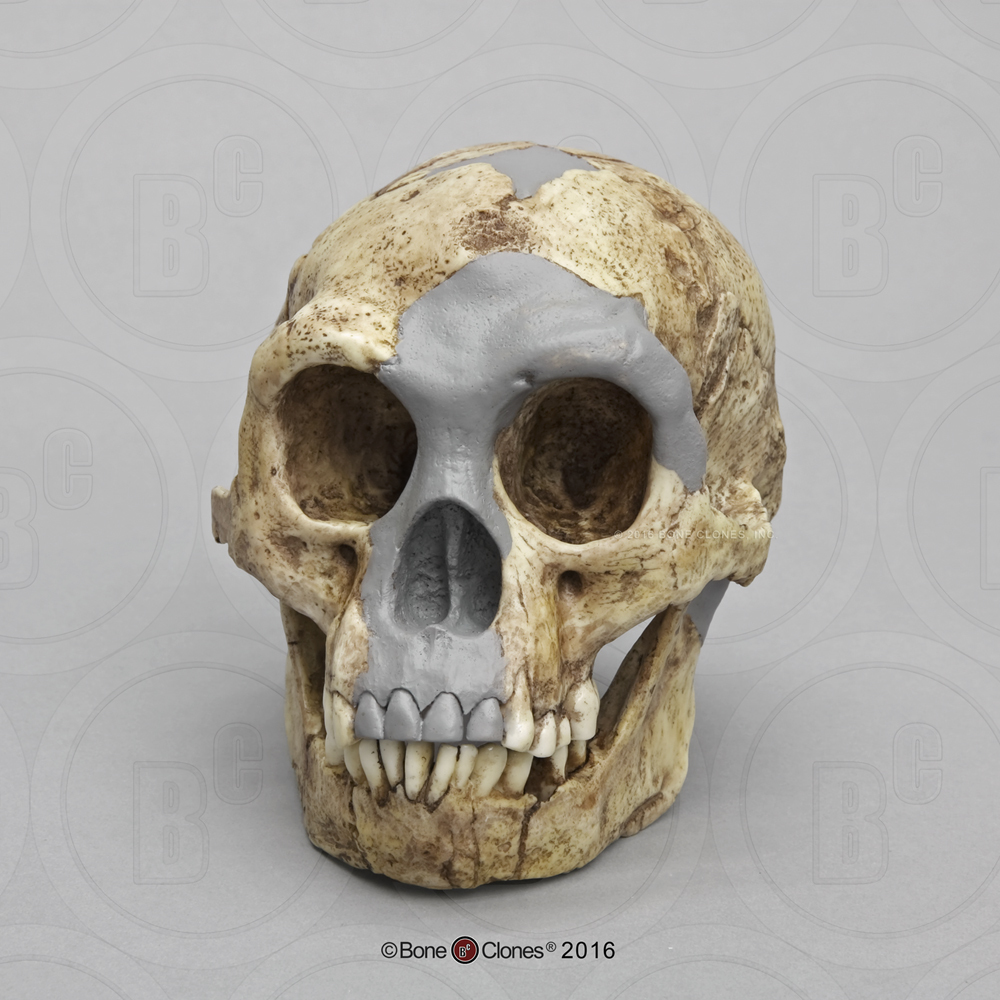
In a small cave called Liang Bua, on the island of Flores, in Indonesia, a small collection of fossils were discovered beginning in 2003 (Figure 7). The fossil fragments represent as many as nine individuals, including a nearly complete female skeleton. The features of the skull are very similar to that of Homo erectus (Figure 8). Homo floresiensis, as the new species is called, had a brain size that was remarkably small at 400 cc, and recent genetic studies suggest a common ancestor with modern humans that predates Homo erectus.
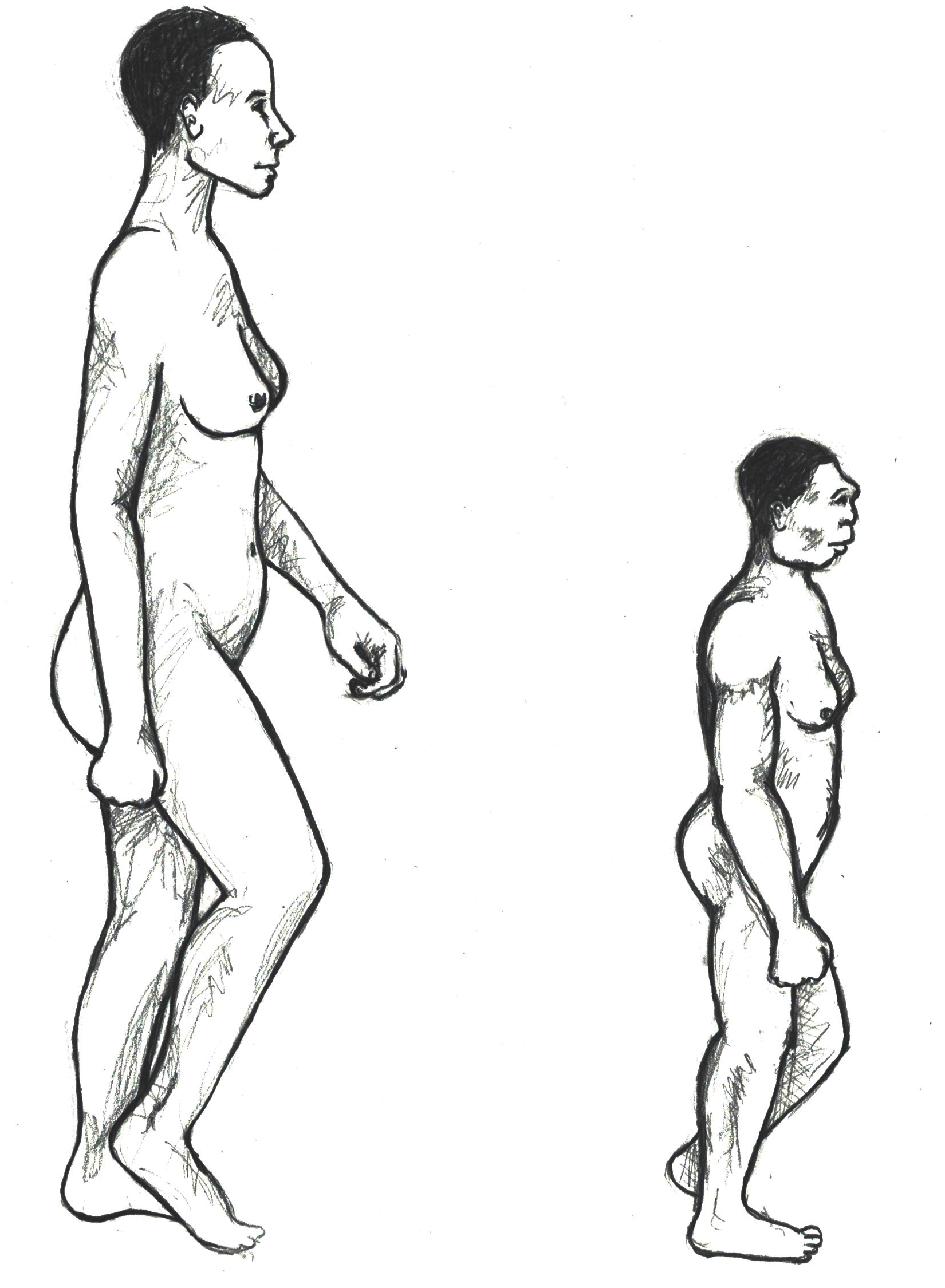
The complete female skeleton, who was an adult, was approximately a meter tall and would have weighed just under 30 kg, which is significantly shorter and just a few kilograms more than the average, modern young elementary-aged child. A reconstructed comparison between an anatomically modern human and Homo floresiensis can be seen in Figure 9. The small size of the fossil has earned the species the nickname “the Hobbit.” Many questions have been asked about the stature of this species, as all of the specimens found also show evidence of diminutive stature and small brain size. Some explanations include pathology; however, this seems increasingly unlikely as all fossils found thus far demonstrate the same pattern. Another possible explanation lies in a biological phenomena seen in other animal species also found on the island and which date to a similar time period. This phenomenon, called insular dwarfing, is due to limited food resources on an island, which can create a selective pressure for large-bodied species to be selected for smaller size, as an island would not have been able to support their larger-bodied cousins for a long period of time. This phenomenon is the cause of other unique species known to have lived on the island at the same time, including the miniature stegadon, a dwarf elephant species.
Homo floresiensis fossils have been dated to have lived on the island between 100,000 and at least 60,000 years ago. There is ongoing research and debate regarding Homo floresiensis’ dates of existence, with some researchers concluding that they lived on Flores until perhaps as recently as 17,000 years ago. Stone tools were also uncovered that have dates overlapping with those of the site and are similar in nature to other hominin stone tools found on the island of Flores. Homo floresiensis would have hunted a wide range of animals, including the miniature stegadon, giant rats, and other large rodents. Other animals on the island that could have threatened them include the giant komodo dragon. An interesting note about this island chain is that ancestors of Homo floresiensis would have had to access the open ocean in order to get there, as the nearest island is almost 10 km away, and there is little evidence to support that a land bridge connecting mainland Asia or Australia to the island would have been present. This would also have limited the number of other animals, including predators as well as human species, that would have had access to the island. Anatomically modern Homo sapiens arrived on the island around 30,000 years ago and may have lived there at the same time as Homo floresiensis, if some researchers’ later dates for Homo floresiensis’ occupation are correct. The modern population living on the island of Flores today believes that their ancestors came from the Liang Bua cave; however, recent genetic studies have determined they are not related to Homo floresiensis.
Homo naledi and Homo floresiensis are clear outliers when compared to their contemporary hominin species. Each has surprised paleoanthropologists for both their archaic traits in relatively modern times and their unique combination of traits seen in archaic species and modern species of humans. While these finds have been exciting, they have also challenged the assumed trajectory of the human lineage, causing scientists to re-examine their previously held assumptions about hominin evolution and what it means to be modern. Add this to the developments being made using ancient DNA, other new fossil discoveries, and other innovations in paleoanthropology, and you see that our understanding of archaic Homo sapiens and others living during this time period is rapidly developing and changing. This is a true testament to the nature of science and the scientific method!
Modern Humans

The walls of a pink limestone cave exposed to the outside world in the hillside of Jebel Irhoud jutted out of the otherwise barren landscape of the Moroccan desert (Figure 10). The year was 2007 and it turned out to be a momentous occasion for science. A fossil unearthed by a team of researchers was barely visible to the untrained eye. Just the fossil cranium’s robust brows were peering out of the rock. The find was welcome but not sheer luck: Hominin fossils have been found here since their first accidental discovery by miners in 1960. This research team from the Max Planck Institute for Evolutionary Anthropology was just the latest to explore the prehistoric human presence in this part of North Africa. Excavating near the first discovery, the researchers wanted to learn more about how Homo sapiens lived far from East Africa, where we thought our species originated.
The scientists were surprised when they analyzed the cranium, named Irhoud 10, and other fossils. Statistical comparisons with other human crania concluded that the Irhoud face shapes were typical of recent modern humans while the braincases matched ancient modern humans. Based on the findings of other scientists, the team expected these modern Homo sapiens fossils to be around 200,000 years old. Instead, dating revealed that the cranium had been buried for around 315,000 years.
Together, the modern-looking facial dimensions and the older date changed the interpretation of our species, modern Homo sapiens. Our key evolutionary changes from archaic Homo sapiens to our species today happened 100,000 years earlier than what we had thought. In addition, the new information suggests that our home region covered more of the vast African continent, instead of being concentrated in the east.
This big addition to the study of modern Homo sapiens is just one of the latest in this continually advancing area of biological anthropology. Researchers are continually discovering amazing fossils and ingenious ways to collect data and test hypotheses about our past. Through the collective work of scientists, including archaeologists, geneticists, and anatomists, we are building an overall theory or explanation of modern human origins. We will first cover the skeletal changes from archaic Homo sapiens to modern Homo sapiens. Next, we will track how modern Homo sapiens expanded the range of its species around the world. Lastly, we will cover the development of agriculture and how it changed human culture to how we practice it today.
Defining Modernity
What defines a modern Homo sapiens when compared to an archaic Homo sapiens? Modern humans, like you and me, have a set of derived traits that are not seen in archaic humans or any other hominin. As with other transitions in hominin evolution, such as increasing brain size and bipedal ability, modern traits do not appear fully formed or all at once. In other words, the first modern Homo sapiens was not just born one day from archaic parents. The traits common to modern Homo sapiens appeared in a mosaic manner: gradually and out of sync with one another. There are two areas to consider when tracking the complex evolution of modern human traits. One is the physical change in the skeleton. The other is behavior inferred from the cranium and material culture.
Skeletal Traits
The skeleton of a modern Homo sapiens is less robust than that of an archaic Homo sapiens. In other words, the modern skeleton is gracile, meaning that the structures are thinner and smoother. Differences related to gracility in the cranium are seen in the braincase, the face, and the mandible. There are also broad differences in the rest of the skeleton.
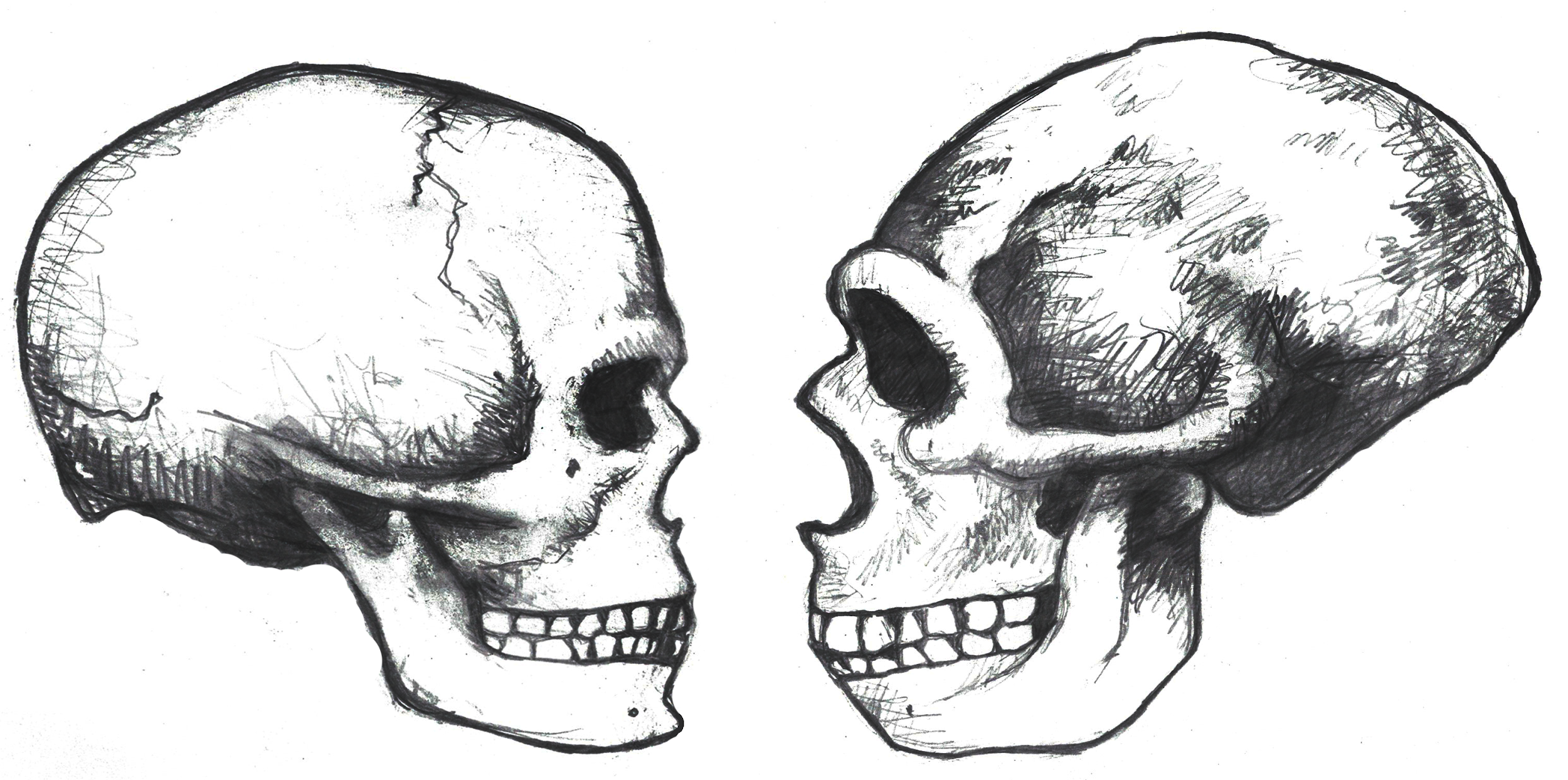
Several elements of the braincase differ between modern and archaic Homo sapiens. Overall, the shape is much rounder, or more globular on a modern skull (Neubauer, Hublin, and Gunz 2018) (Figure 11). Viewed from the side, the tall vertical forehead of a modern Homo sapiens stands out when compared to the sloping archaic version. This is because the frontal lobe of the modern human brain is larger than the one in archaic humans, and the skull has to accommodate the expansion. The different priorities in brain regions may also indicate cognitive and behavioral differences between archaic humans and modern humans.
The trend of shrinking face size across hominins reaches its extreme with our species as well. The facial bones of a modern Homo sapiens are extremely gracile compared to all other hominins (Lieberman, McBratney, and Krovitz 2002). As with the shrinking of the face leading up to our species, the decreasing reliance on needing large teeth for survival may have been the reason that modern human faces are so gracile in comparison to other humans. Continuing a trend in hominin evolution, technological innovations kept reducing the importance of teeth in reproductive success. As natural selection favored smaller and smaller teeth, the surrounding bone holding these teeth also shrank.
Connected to the face, the mandible is also gracile in modern humans when compared to archaic humans and other hominins. Interestingly, our mandibles have pulled back so far from the prognathism of earlier hominins that we gained an extra structure at the most anterior point, called the mental eminence. You know this structure as the chin. The chin acts as structural support to withstand strain on the otherwise gracile mandible. In other words, as natural selection favored smaller dentition, the chin developed to maintain structural integrity of the mandible.
Overall, a modern Homo sapiens post-cranial skeleton has thinner bone, smoother features, and more slender shapes when compared to archaic Homo sapiens (Figure 12). Comparing whole skeletons, modern humans have longer limb proportions relative to the length and width of the torso, giving us lankier outlines.
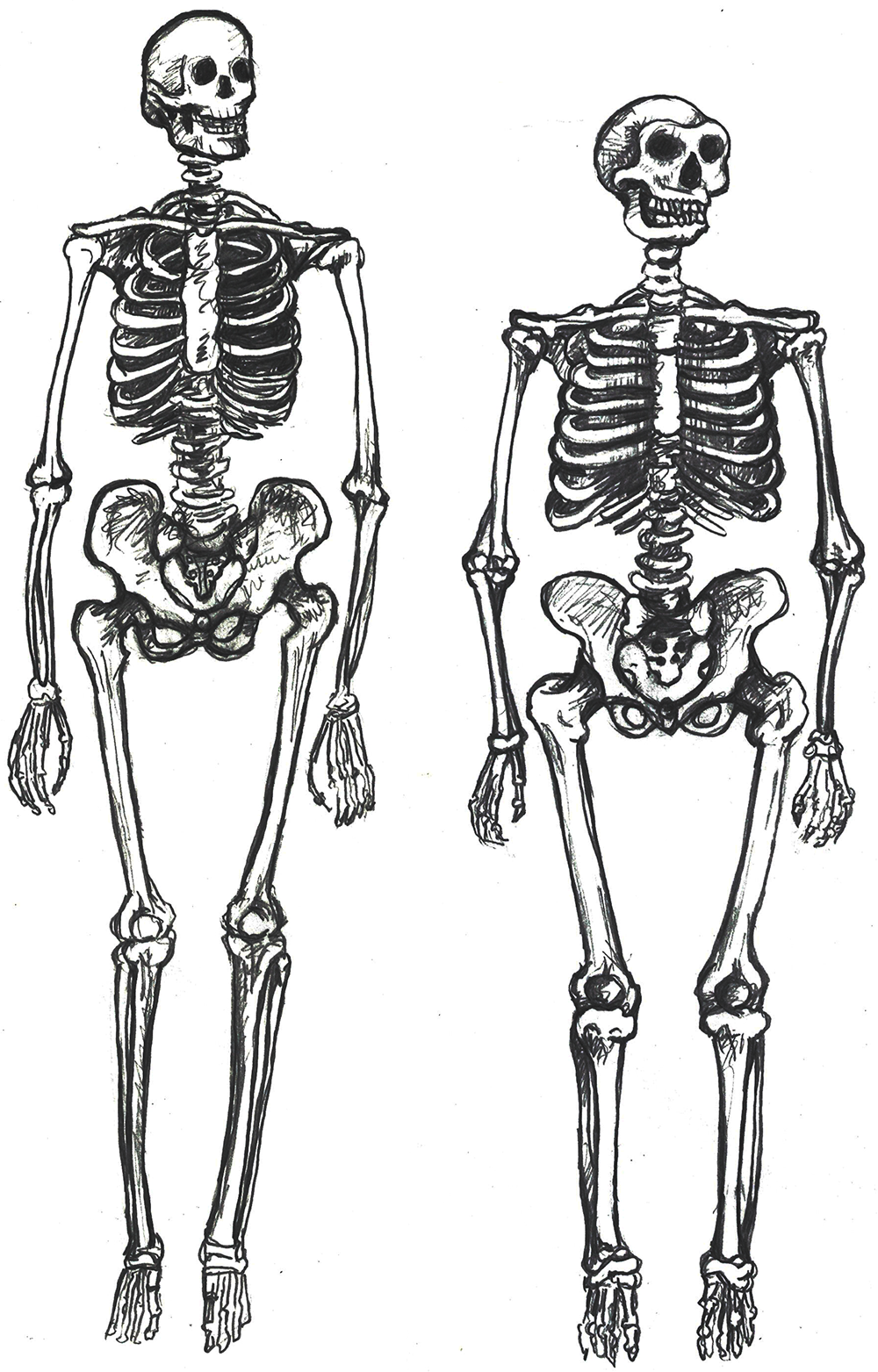
A slender frame is adapted for the efficient long-distance running ability that started with Homo erectus. Furthermore, slenderness is a genetic adaptation for cooling an active body in hotter climates, which aligns with the ample evidence that Africa was the home continent of our species.
Behavioral Modernity
Aside from physical differences in the skeleton, researchers have also tracked clues of behavioral changes from archaic to modern humans. From the anthropology of our species today, we know that we practice a very complex version of culture, with many layers to our language, art, social organization, and technology, among other areas. Did cultural complexity increase gradually or quickly with the first modern humans? This question is being actively investigated. A major obstacle to answering this question is that it is hard to define and measure cultural complexity. Since we cannot directly observe humans of the distant past, we have to infer these measures of human behavior from other types of evidence. Two particularly illuminating areas are archaeology and the analysis of reconstructed brains.
Archaeology tells us much about the behavioral complexity of past humans by interpreting the significance of material culture. In terms of evolved advanced culture, items created with an artistic flair, or as a decorative piece, speak of abstract thought processes (Figure 13). The demonstration of difficult artistic techniques and technological skills hints at social learning and cooperation as well. For example, most of your skills were taught to you by a more experienced person, upon which you’ve developed your own style with practice. Some day you may pass on what you know to someone else using language to convey your knowledge. The same process is believed to have happened with early modern humans in areas such as toolmaking and craftwork, producing the sophisticated material culture that we can now study. According to paleoanthropologist John Shea (2011), one way to track the complexity of past behavior through artifacts is by measuring the variety of tools found together. The more types of tools constructed with different techniques and for different purposes, the more modern the behavior. Turning this view to ourselves, think of all of the tools we have available to us today at a typical hardware store and the cumulative knowledge they represent. This idea of measuring past behavior is promising, but researchers are still working on an archaeological way to measure cultural complexity that is useful across time and place.

The interpretation of brain anatomy is another promising approach to studying the evolution of human behavior. When looking at the body of work on this topic in modern Homo sapiens brains, researchers found a weak association between brain size and test-measured intelligence (Pietschnig et al. 2015). This means that there are more significant factors that affect tested intelligence than just brain size. Additionally, they found no association between intelligence and biological sex. Since the sheer size of the brain is not useful for weighing intelligence, paleoanthropologists are instead investigating the differences in certain brain structures. The differences in organization between modern Homo sapiens brains and archaic Homo sapiens brains may reflect different cognitive priorities that account for modern human culture. Researchers have hypothesized that the expanded frontal and parietal lobes in the globular modern human braincases mean that we can do more complex thinking regarding memory and social ability than the Neanderthals could. In contrast, the Neanderthal brain prioritized the visual regions where the occipital bun was located, with fewer neurons in the frontal area for complex thinking. As with the archaeological line of research in the preceding paragraph, this is a very active area of investigation. New discoveries will refine what we know about the human brain and apply that knowledge to studying the distant past.
Taken together, the cognitive abilities in modern humans may have translated into an adept use of tools to enhance survival. The ability to process a new environment, adapt to it with innovative technology, and pass on that knowledge may be the key behind the success of modern Homo sapiens.
First Africa, Then the World
What enabled modern Homo sapiens to expand its range further in 300,000 years than Homo erectus did in 1.5 million years? The key is the set of derived biological traits discussed in the last section. The gracile frame and neurological anatomy allowed modern humans to survive and even flourish in the vastly different environments they encountered. Based on multiple types of evidence, the source of all of these modern humans, including all of us today, was Africa.
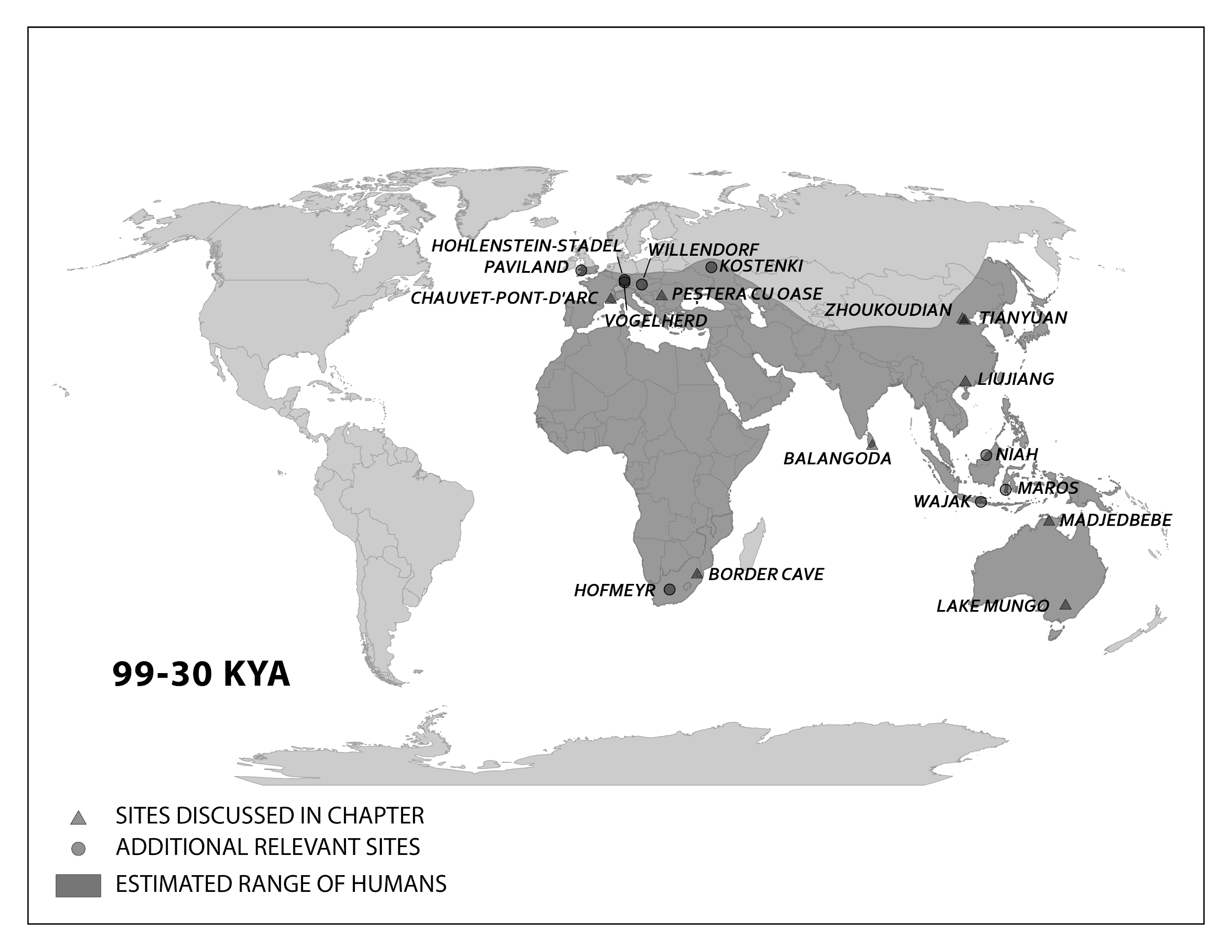
This section traces the origin of modern Homo sapiens and the massive expansion of our species across all of the continents except Antarctica by 12,000 years ago. While modern Homo sapiens first shared geography with archaic humans, modern humans eventually spread into lands where no human had gone before (Figure 14). Starting with the first-known modern Homo sapiens, around 315,000 years ago, we will follow our species from a time called the Middle Pleistocene to the end of the Late Pleistocene. Culturally, we will trace developments from the Middle Stone Age through the transition around 50,000 years ago to the Later Stone Age.
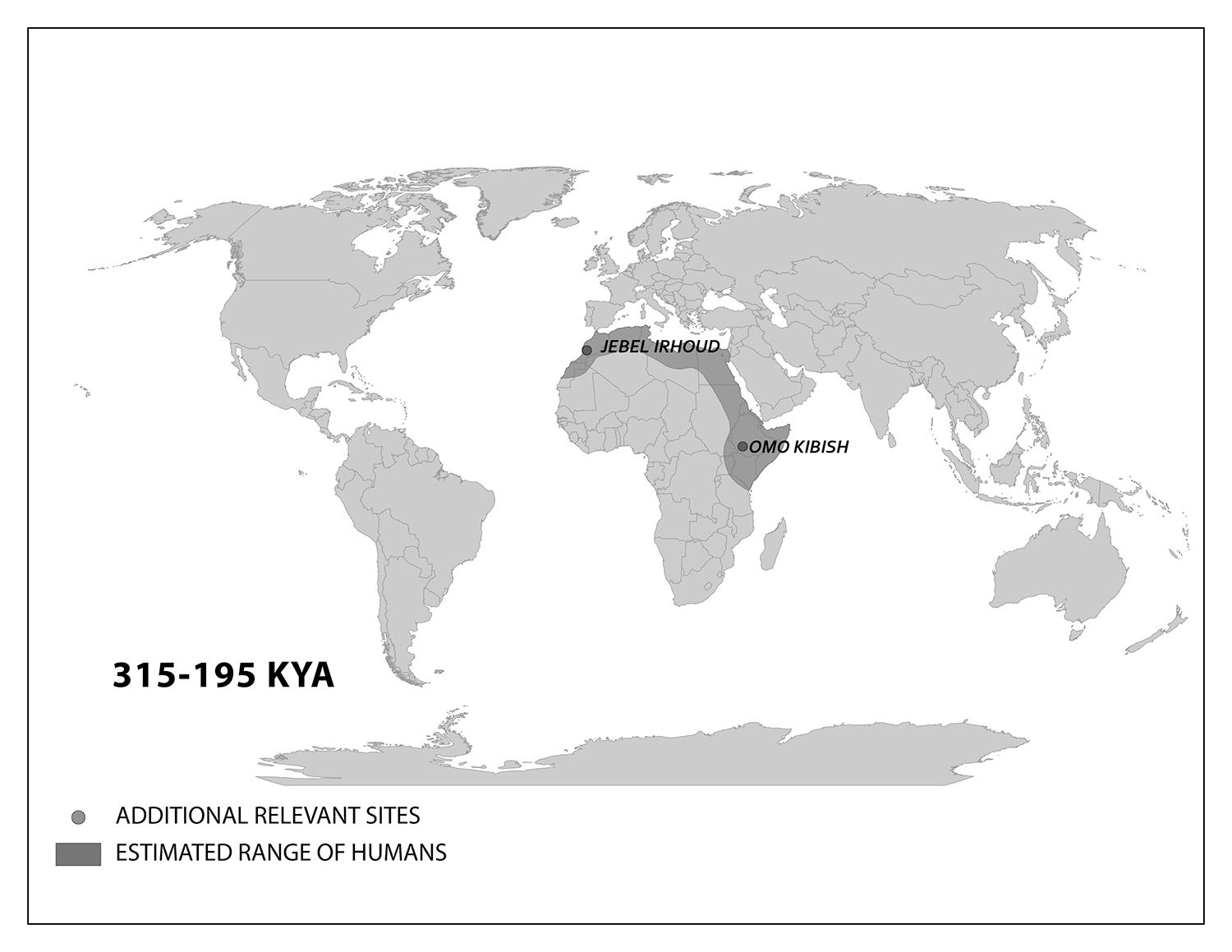
The Start of Modern Homo sapiens in Africa
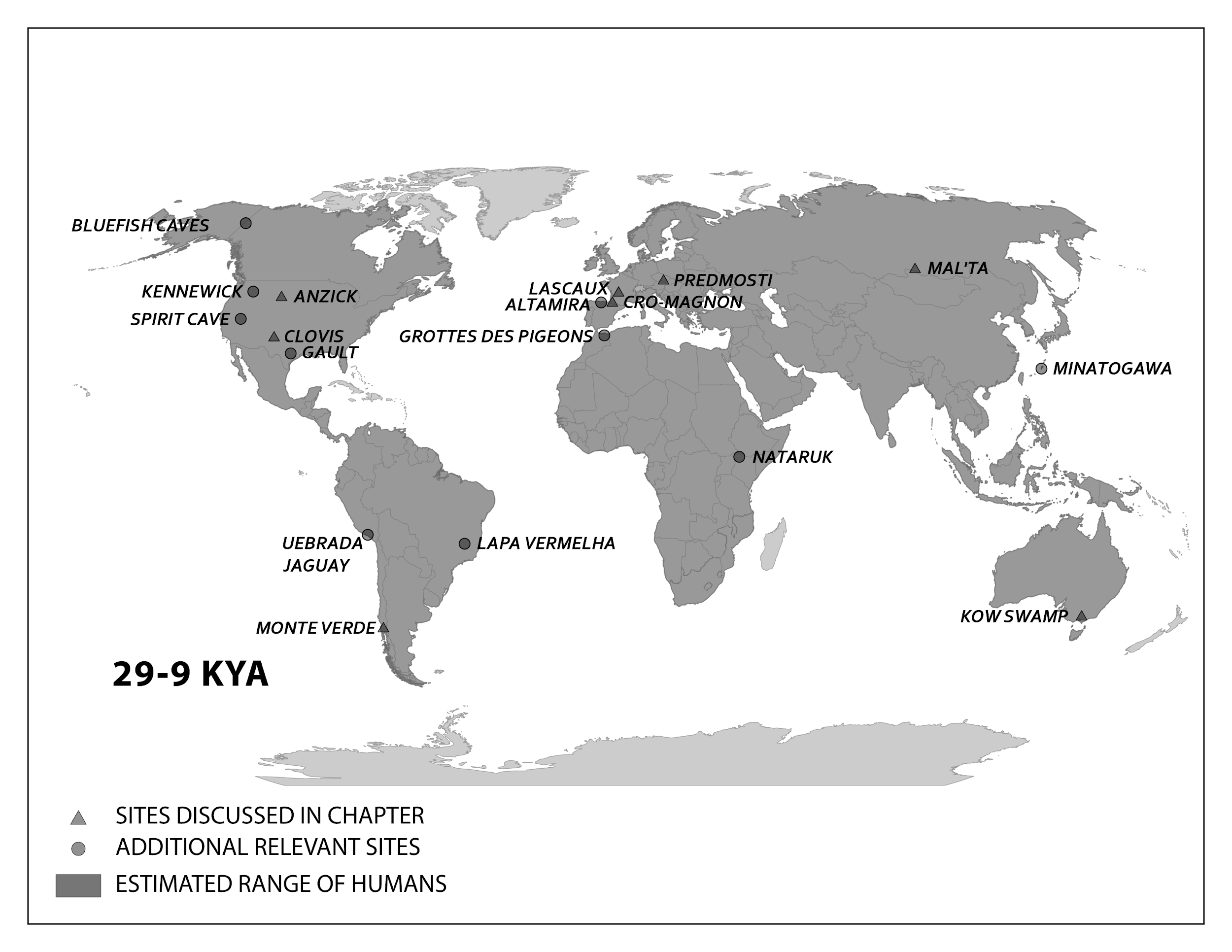
We start with the ample fossil evidence supporting the theory that modern humans originated in Africa during the Middle Pleistocene, having evolved from African archaic Homo sapiens. The earliest dated fossils considered to be modern actually have a mosaic of archaic and modern traits, showing the complex changes from one type to the other. Experts have various names for these transitional fossils, such as ‘Early Modern Homo sapiens’ or ‘Early Anatomically Modern Humans.’ However they are labeled, the presence of some modern traits means that they illustrate the origin of the modern type. Three particularly informative sites with fossils of the earliest modern Homo sapiens are Jebel Irhoud, Omo, and Herto.
Recall from the start of the chapter that the most recent finds at Jebel Irhoud are now the oldest dated fossils that exhibit the traits of modern Homo sapiens. Besides Irhoud 10, the cranium that was dated to 315,000 years ago (Hublin et al. 2017), there were other fossils found in the same deposit that we now know are from the same time period. In total there are at least five individuals, representing life stages from childhood to adulthood (Figure 15)
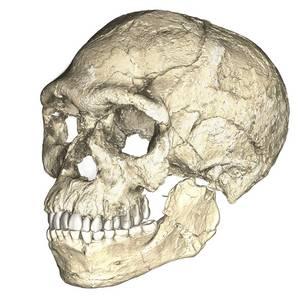
Research separated by nearly four decades uncovered fossils and artifacts from the Kibish Formation in the Lower Omo Valley in Ethiopia. These Omo Kibish hominins were represented by braincases and fragmented postcranial bones of three individuals found kilometers apart, dating back to 195,000 years ago (McDougall, Brown, and Fleagle 2005).
Also in Ethiopia, a team led by Tim White (2003) excavated numerous fossils at Herto. There were fossilized crania of two adults (Figure 16) and a child, along with fragments of more individuals. The dates ranged between 160,000 and 154,000 years ago. The skeletal traits and stone tool assemblage were both intermediate between the archaic and modern types.

The combined fossil evidence paints a picture of diversity in geography and traits. Instead of evolving in just East Africa, the Jebel Irhoud find revealed that early modern Homo sapiens had a wide range across Middle Pleistocene Africa. The hypothesis that there was no single original home within Africa for our species is called African multiregionalism (Scerri et al. 2018). Supporting this explanation, fossils have different mosaics of archaic and modern traits in different places and even within the same area. The high level of diversity from just these fossils shows that the modern traits took separate paths toward the set we have today.
What about behavioral modernity? Jebel Irhoud, Omo, and Herto all bore Middle Stone Age tools of the same flaked style as archaic assemblages, even though they were separated by almost 150,000 years. No clear signs of art dating back this far have been found either. Other hypotheses not related to behavioral modernity could explain these observations. The tool set may have been suitable for thriving in Africa without further innovation. As for the lack of art, maybe works from that time were made with media that deteriorated or perhaps such works were removed by later humans.
While modern Homo sapiens lived across Africa, some members eventually left the continent. Generations of these pioneers entered environments far different from what their ancestors experienced in Africa.
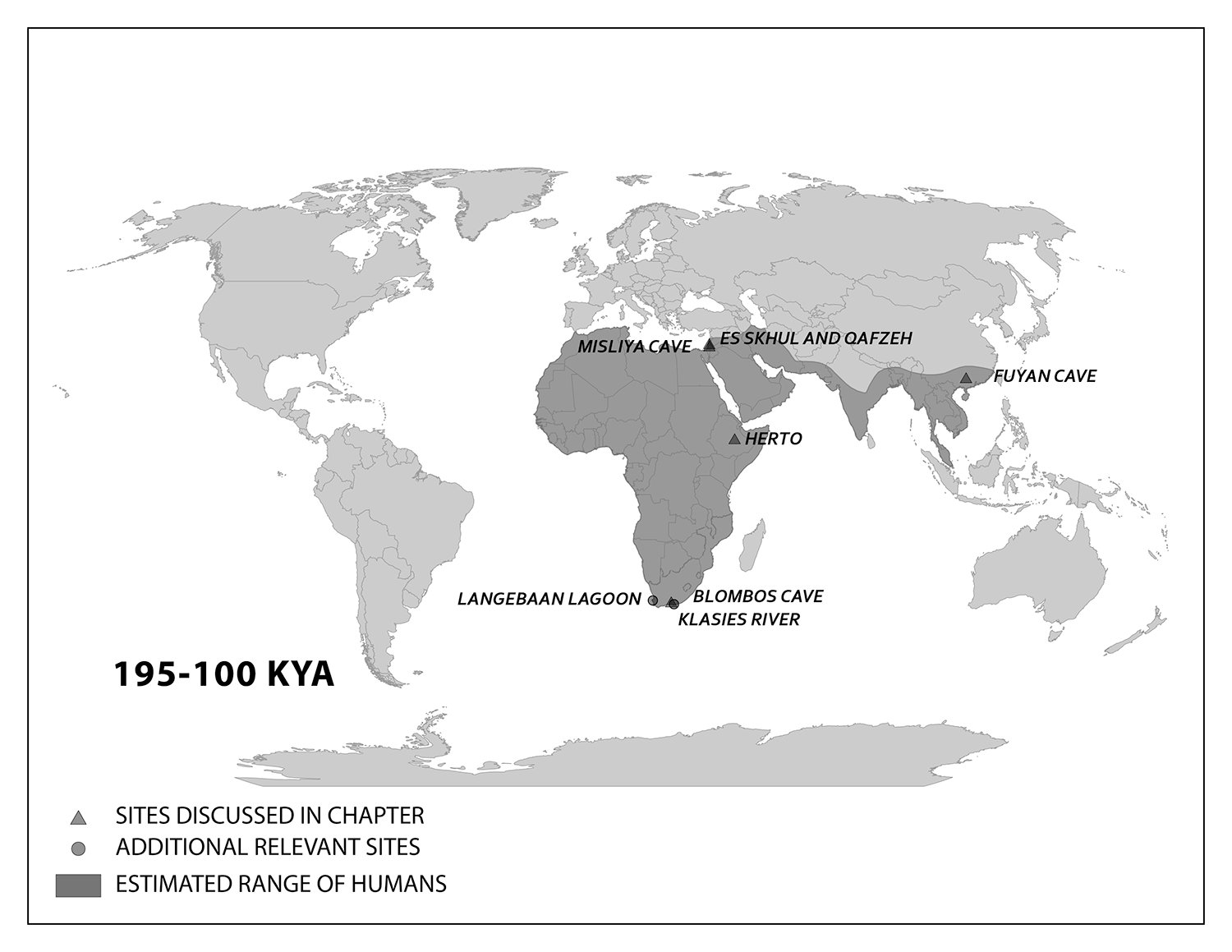
Expansion into the Middle East and Asia
This section presents key finds showing where modern Homo sapiens went after the range of the species first extended out of Africa. These pioneers could have used several connections to the Middle East, or West Asia. From North Africa, they could have crossed the Sinai Peninsula and moved north to the Levant, or eastern Mediterranean. Finds in that region show an early modern human presence. There were likely multiple waves of movement producing gene flow back and forth across these regions. The expanding modern human population could have thrived by using resources along the southern coast of the Arabian Peninsula to South Asia, with side routes moving north along rivers.
Geographically, the Middle East is the ideal place for the African modern Homo sapiens population to inhabit upon expanding out of their home continent. In the Eastern Mediterranean coast of the Levant, there is a wealth of skeletal and material culture linked to modern Homo sapiens. Recent discoveries from Saudi Arabia further add to our view of human life just beyond Africa.
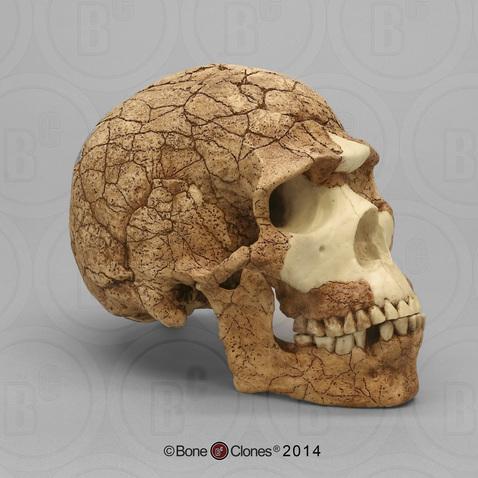
The Caves of Mount Carmel in present-day Israel have preserved skeletal remains and artifacts of modern Homo sapiens, the first-known group living outside Africa. From 120,000 to 90,000 years ago, fossils of multiple individuals across life stages were found in the caves of Es-Skhul and Qafzeh (Shea and Bar-Yosef 2005). The skeletons had many modern Homo sapiens traits, such as globular crania and more gracile postcranial bones when compared to Neanderthals. Still, Skhul V had a particularly impressive brow ridge that was short in height but sharply jutted forward above the eyes (Figure 17). The high level of preservation is due to the intentional burial of some of these people. Besides skeletal material, there are signs of artistic or symbolic behavior. For example, the adult male Skhul V had a boar’s jaw on his chest. Similarly, Qafzeh 11, a juvenile with healed cranial trauma, had an impressive deer antler rack placed over his torso (Figure 18). Perforated seashells colored with ochre were also found in Qafzeh.

One remaining question is, what happened to the modern humans of the Levant after 90,000 years ago? Another site attributed to our species did not appear in the region until 47,000 years ago. Competition with Neanderthals may have accounted for the disappearance of modern human occupation since the Neanderthal presence in the Levant lasted longer than the dates of the early modern Homo sapiens.
A long history of paleoanthropology in China has found ample evidence of modern human presence. Four notable sites are the caves at Fuyan, Liujiang, Tianyuan, and Zhoukoudian. In the distant past, these caves would have been at least seasonal shelters that unintentionally preserved evidence of human presence for modern researchers to discover.
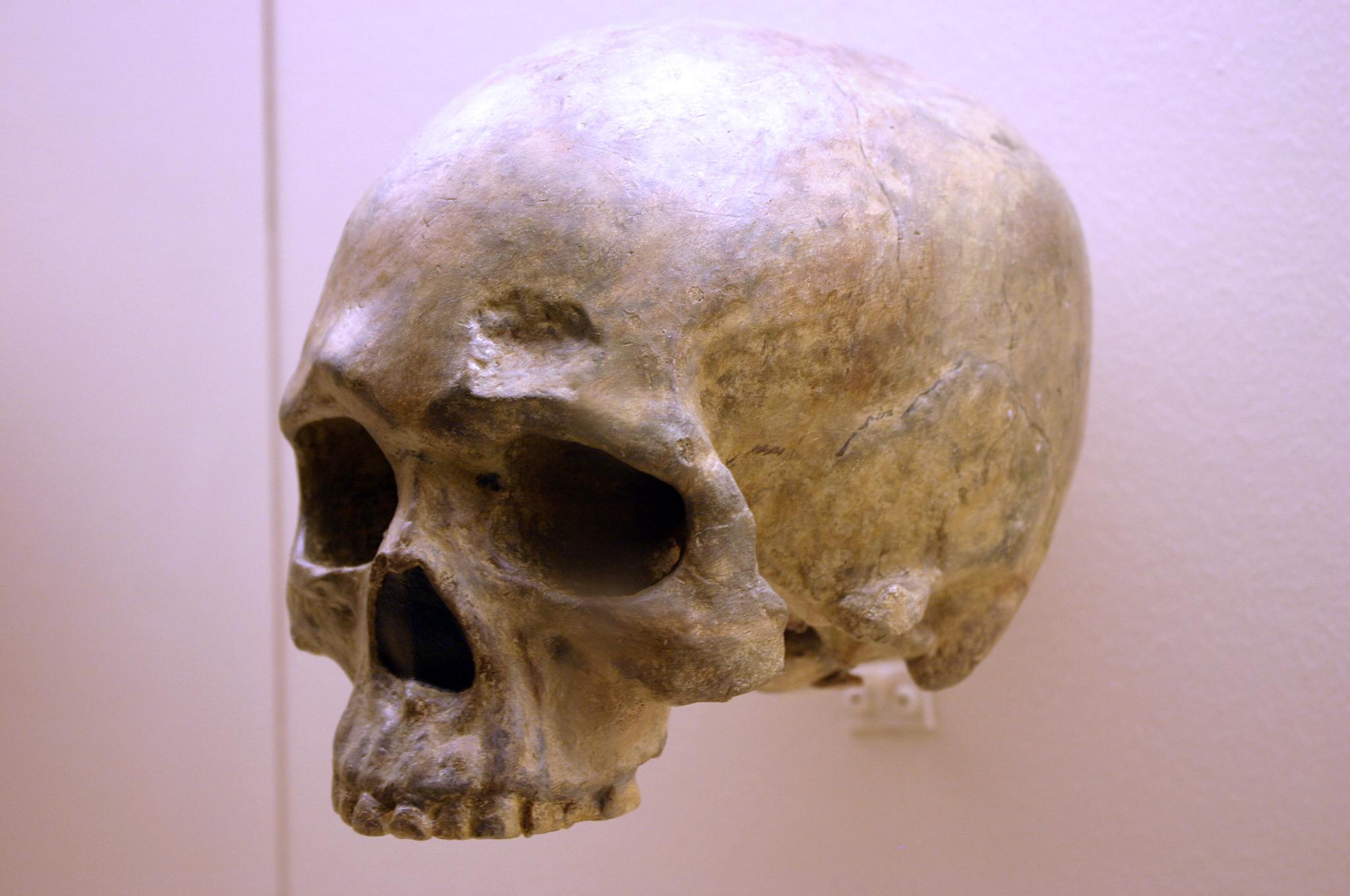
At Fuyan Cave in Southern China, paleoanthropologists found 47 adult teeth associated with cave formations dated to between 120,000 and 80,000 years ago (Liu et al. 2015). It is currently the oldest-known modern human site in China, though other researchers question the validity of the date range. The teeth have the small size and gracile features of modern Homo sapiens dentition. No lithics have been found in Fuyan Cave. The fossil Liujiang (or Liukiang) hominin has derived traits that classified it as a modern Homo sapiens, though primitive archaic traits were also present (Figure 19). Dating methods suggested an age of around 67,000 years. A mandible fragment, teeth, and postcranial skeletal remains of a single adult of indeterminate sex was found by tree farmers in Tianyuan, 50 km from Beijing. Radiocarbon dating of the bones estimated that they were from 42,000 to 39,000 years ago.
The last Chinese site to describe here is the one that has been studied the longest. In the Zhoukoudian Cave system, where Homo erectus and archaic Homo sapiens have also been found, there were three crania that fit the modern Homo sapiens set of traits (Figure 7.11). These crania were in a part of the cave called the Upper Cave, dating to between 34,000 and 10,000 years ago. Comparison of cranial measurements to other populations past and present found no connection with modern East Asians. These findings again show that human variation was very different from what we see today.
As in Africa, the finds of the Middle East have shown that humans were biologically diverse and had complex relationships with their environment. Work in the Levant showed an initial expansion north from the Sinai Peninsula that did not last. Local resources were used to make lithics and decorative items. The early Asian presence of modern Homo sapiens was complex and varied as befitting the massive continent. What the evidence shows is that people adapted to a wide array of environments that were far removed from Africa. Faunal and floral remains found in these shelters speak to the flexibility of the human omnivorous diet as local wildlife and foliage became nourishment. Decorative items, often found as burial goods in planned graves, show a flourishing cultural life. Eventually, modern humans at the southeastern fringe of the geographical range of the species found their way southeast until some became the first humans in Australia.
Crossing to Australia
Expansion of the first modern human Asians, still following the coast, eventually entered an area called Sunda by researchers before continuing on to modern Australia. Sunda was a landmass made up of the modern-day Malay Peninsula, Sumatra, Java, and Borneo. Lowered sea levels connected these places with land bridges, making them easier to traverse. Proceeding past Sunda meant navigating Wallacea, the archipelago that includes the Indonesian islands east of Borneo. Prehistorically, there were many megafauna, large animals that migrating humans would have used for food and materials such as hides and bones. Further southeast was another prehistoric landmass called Sahul, which included New Guinea and Australia as one contiguous continent. This land had never seen hominins or any other primates before modern Homo sapiens arrived. Sites along this path offer clues about how our species handled these changes to the local environment to live successfully as foragers.
The skeletal remains at Lake Mungo are the oldest known in Australia. The lake, now dry, was one of a series located along the southern coast of Australia in New South Wales, far from where the first people entered from the north. Two individuals dating to around 40,000 years ago show signs of artistic and symbolic behavior, including intentional burial. The bones of Lake Mungo 1 (LM1), an adult female, were crushed repeatedly, colored with red ochre, and even cremated (Bowler et al. 1970).
Kow Swamp, also in southern Australia, contained human crania that looked distinctly different from the ones at Lake Mungo. The Kow Swamp crania had extremely robust brow ridges and thick bone walls, but these were paired with globular features on the braincase (Figure 20). The frontal bones had extremely linear slopes from the brow to the top of the cranium, resembling intentional cranial modification seen in other parts of the world. If the crania were shaped on purpose, they are another sign of symbolic behavior, as the practice has linked to ideas of group cultural identity. By the time of the Kow Swamp people, between 9,000 and 20,000 years ago, cranial modification may have been a meaningful part of culture in southern Australia.

The presence of the first humans in Australia along the current northern and southern coasts suggests that they used a route that wrapped around the perimeter of the continent. This path allowed access to both coastal and inland resources. Megafauna was a likely source of food and other resources. The mythology of Australian aborigines today has been linked by researchers to extinct life, such as marsupial tapirs and lions. Predation by humans may be why the megafauna became extinct, leaving the oral tradition of their existence.
The abundant evidence matching the criteria for behavioral modernity shows that the early Australians had a rich artistic and symbolic life. Raw materials must have been transported or traded across long distances in order to make art and color both human and nonhuman skeletal remains. The local varieties of stone tools and art may reflect cultural variation across distant regions of the continent.
Northwest to Europe
The first modern human expansion into Europe occurred after other members of our species settled East Asia and Australia. As the evidence from the Levant suggests, modern human movement to Europe may have been hampered by the presence of Neanderthals. Another obstacle was that the colder climate was incompatible with the biology of African modern Homo sapiens, which was adapted for exposure to high heat and ultraviolet radiation. Still, by 40,000 years ago, modern Homo sapiens had enough of a presence in Europe to leave evidence for researchers to find. This time was also the start of the Later Stone Age. Connected with the history of science in general, early modern Homo sapiens in Europe have been studied for centuries. Due to the bias in research focus favoring Europe, there is a wealth of evidence to explore.
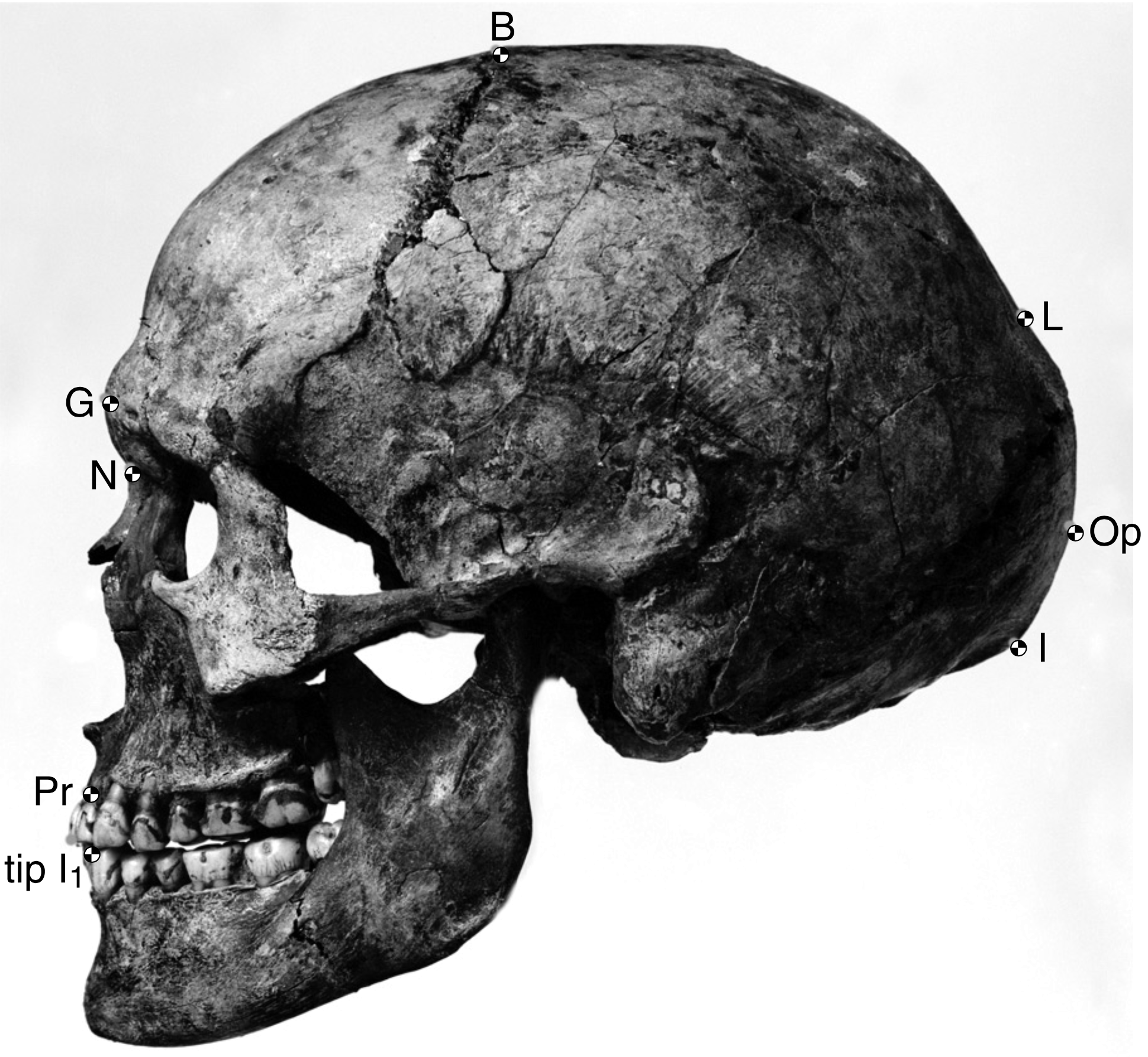
In Romania, the site of Peștera cu Oase (Cave of Bones) had the oldest known remains of modern Homo sapiens in Europe, dated to around 40,000 years ago (Trinkaus et al. 2003). Among the bones and teeth of cave bears, wolves, ibex, and other animals were the fragmented cranium of one person and the mandible of another (the two bones did not fit each other). The DNA from Oase 1 is equally similar to DNA from present-day Europeans and Asians. This means that Oase 1 was not the direct ancestor of modern Europeans.
The term “Cro-Magnon” has entered public usage as a name for any prehistoric modern European Homo sapiens, and maybe any “caveman” of our species, but it technically refers to four adults (three male and one female) and an infant found in the Cro-Magnon rock shelter in France in 1868. The remains are dated to 28,000 years ago and may all have been intentionally buried along with over 300 pierced seashells and nonhuman skeletal remains. The Cro-Magnon crania are easily identifiable by their rectangular eye orbits, which are more angular than any contemporary (Figure 21). Compared to Neanderthal skeletons of the same region, the Cro-Magnons are extremely gracile. The adults also show signs of much pathology, including fused neck vertebrae and healed fractures. The individual Cro-Magnon 1 has skeletal lesions typical of neurofibromatosis type 1, a rare genetic disease that causes tumor growth. The combination of disease markers suggest that life for the Cro-Magnons was so physically demanding that it greatly affected the skeleton.
Dating to around 26,000 years ago, Předmostí near Přerov in the Czech Republic was a site where people buried over 30 individuals along with many artifacts. Eighteen individuals were found in one mass burial area, a few covered by the scapulae of woolly mammoths . While the recovered human skeletons were destroyed in World War II, finely detailed photographic negatives allowed comparisons to other human groups (Figure 22). Besides the human remains, the site contained the bones of over a thousand mammoths. Some of the mammoth remains were shaped by humans, including a limb bone fragment with a carved abstract female figure. There is also skeletal evidence of dog domestication, such as the presence of dog skulls with shorter snouts than in wild wolves. In total, Předmostí could have been a settlement dependent on mammoths for subsistence with people participating in artistic behaviors and the artificial selection of early domesticated dogs.
Among the many European sites dating to the Later Stone Age, the famous cave art sites deserve mention. Chauvet-Pont-d’Arc Cave in southern France dates to separate Aurignacian occupations 31,000 years ago and 26,000 years ago. Over a hundred art pieces representing 13 animal species are preserved. Some depicted species are common to European cave art, such as deer and horses. Others are rare, such as rhinos and owls. Two possible human figures are in the deepest gallery of the cave system. Besides the painted figures, the tracks and skulls of cave bears and an ibex were also found in the cave. Another famous French cave with art is Lascaux, which is several thousand years younger at 17,000 years ago in the Magdalenian period. At this site, there are over 6,000 painted figures on the walls and ceiling (Figure 23). The paint was made of a mix of mineral pigments in liquid binder made from fat or clay. Scaffolding and lighting must have been used to make the paintings on the walls and ceiling deep in the cave. Overall, visiting Lascaux as a contemporary must have been an awesome experience: trekking deeper in the cave lit only by torches giving glimpses of animals all around as mysterious sounds echoed through the galleries. The professionally lit photographs of today do not give the original context justice, though replicas have been built to simulate the experience for tourists. Both Chauvet and Lascaux have been closed to all but researchers due to the degradation of the art when tourism was allowed.

Study of Europe in the Upper Paleolithic gives a more detailed view of the general pattern of biological and cultural change linked with the arrival of modern Homo sapiens. The modern humans experienced a rapidly changing culture that from our perspective went through four major growths in complexity and refinement. The cave art sites showed a deeper use of expression and symbolism, though the exact meaning is unclear. With survival dependent on the surrounding ecology, painting the figures may have connected people to important and impressive wildlife at both a physical and spiritual level. Both reverence for animals and the use of caves for an enhanced sensory experience are common to cultures today and through recorded history.
Distant Relations: Interbreeding with Archaic Humans Outside Africa
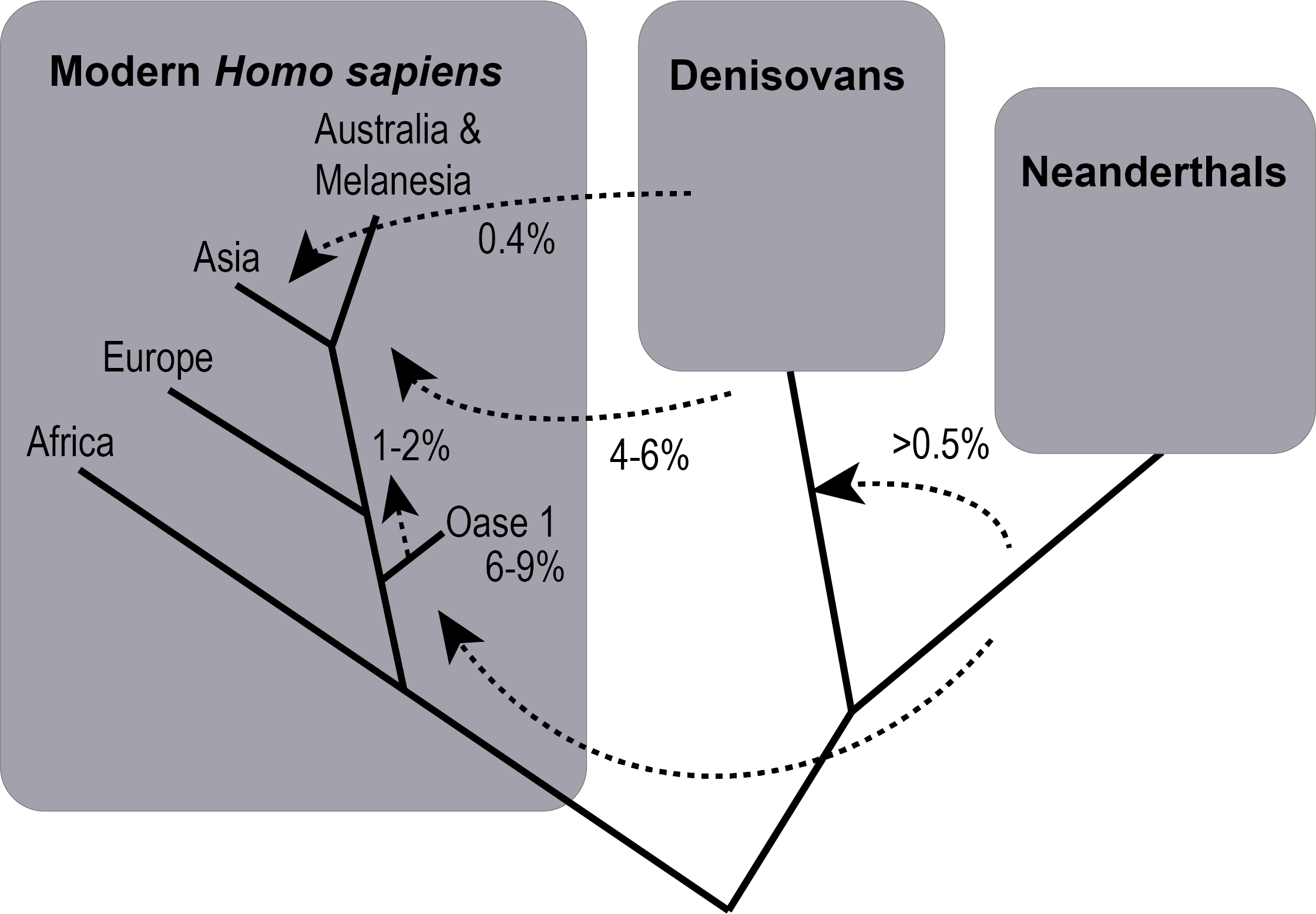
As the modern human population grew beyond Africa, they interbred with the archaic Homo sapiens who were already there, descendants of the Homo erectus populations before them. This statement does not mean that modern humans are the direct descendants of the archaic Neanderthals. Instead, the building evidence suggests a more complex connection between archaic and modern humans outside of Africa. Three Homo sapiens groups interbred with one another: modern Homo sapiens and two archaic groups, the Neanderthals and the Denisovans (Figure 24).
Interbreeding with Neanderthals
Since the first finds of Neanderthal remains, researchers have sought evidence of a biological connection with modern Europeans. While there are no sites that show Neanderthals and modern Homo sapiens lived together (such as both types of skeletons found together in a common burial), interbreeding has been suggested by the appearance of Neanderthal traits in otherwise modern human skeletons found in their common geographical range.
A new source of evidence arrived with the invention of ancient DNA (aDNA) analysis, which has built up more evidence of Neanderthal and modern human interbreeding. DNA samples from Neanderthal fossils have been compared to DNA from both prehistoric and present-day modern Homo sapiens to trace the amount of gene flow between these groups. The amount of transfer is more indicative of introgression, the entrance of small, uneven portions of Neanderthal DNA into modern humans, rather than an even hybridization over time (Dannemann and Racimo 2018). The introgression could have been caused by an imbalance in population size: the continually growing modern population with gene flow from Africa could have diluted the incoming Neanderthal DNA to the low percentage seen today.
The findings of Neanderthal DNA analysis and comparison with modern human genomes worldwide have revealed surprising details about the interactions between these groups. An unexpected result is that modern Asians have more DNA from Neanderthals than modern Europeans despite the separation in geography (Wall and Yoshihara Caldeira Brandt 2016). Today, non-Africans have around 1%–2% Neanderthal DNA in their genomes, with Asians having more than Europeans.
While the study of skeletal traits suggested archaic-modern interbreeding, the use of DNA analysis provided solid evidence of these events between archaic and modern Homo sapiens in Europe and Asia. This is a very active field as the technology keeps improving and more samples are analyzed. A consistent picture is forming, but many of the details concerning the timing and context of interbreeding are still unclear. At this point, we know that three human groups interbred with each other at different times and places. DNA originating from Neanderthals and Denisovans was found in prehistoric modern humans and is present in some Europeans and Asians of today. The introgressed DNA has affected the traits of our own species and they are just being discovered.
African Developments
We now switch our view back to Africa to see what developments occurred after members of our species first crossed to the Middle East and beyond. Our survey of modern Homo sapiens expansion left Africa around 150,000 years ago to see where people pushed the fringes of our geographical range. It is important to remember the species did not all leave Africa together. While modern humans found ways to survive in the Middle East, Asia, Sahul, and Europe, many others remained in Africa. Evidence of what Homo sapiens did in Africa from the end of the Middle Stone Age to the Later Stone Age is concentrated in South African sites. There, Blombos Cave and Border Cave show that complex human behavior was developing in the home continent, with the earliest evidence of many cultural practices.
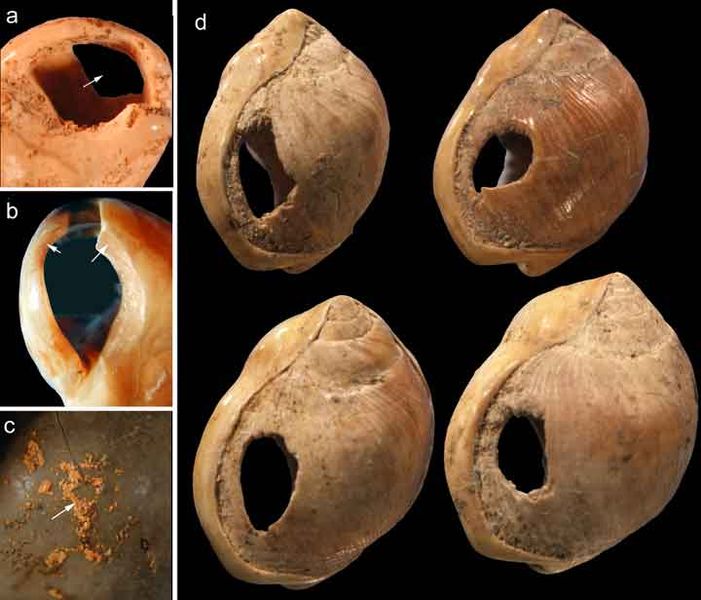
Blombos Cave is located along the present shore of the Cape of Africa facing the Indian Ocean and is notable for having a wide variety of artifacts. The material culture shows that toolmaking and artistry were more complex than previously thought for the Middle Stone Age. Excavations and analysis have been carried out since the 1990s by a research team including Francesco d’Errico, Christopher Henshilwood, and Marian Vanhaeren. In a layer dated to 100,000 years ago, researchers found two intact ochre-processing kits made of abalone shells and grinding stones (Henshilwood et al. 2011). Chemical analysis found the likely ingredients of the ochre-based paint, including crushed bone, animal fat, and charcoal. More ochre fragments, including some marked with notches, were found all throughout the site. The team performed numerous analyses and experiments to show that perforated marine snail shell beads from 75,000 years ago were shaped by people using bone points found in the cave (Figure 25). Together, the evidence shows that the Middle Stone Age occupation at Blombos Cave incorporated resources from a variety of local environments into their culture, from caves (ochre), open land (animal bones and fat), and the sea (abalone and snail shells). This complexity shows a deep knowledge of the region’s resources and their use—not just for survival but also for symbolic purposes.
On the eastern coast of South Africa, Border Cave shows new African cultural developments at the start of the Later Stone Age. Paola Villa and colleagues (2012) identified several changes in technology around 43,000 years ago. Stone tool production transitioned from a slower, measured process to one that took less time to finish but made many microliths. An adhesive made from tree bark was found on some of the microliths. The researchers hypothesize that hunting technology moved from large crafted spearheads to smaller bone arrow points that were adhered to shafts and even tipped with poison for more effectiveness. Changes in decorations were also found across the Later Stone Age transition. Beads were made from a new resource: fragments of ostrich eggs. Unlike the snail shell beads, which retained the shape of the original structure, ostrich shell beads were shaped into circular forms, resembling present-day breakfast cereal O’s. While a subtle difference, these beads show a higher level of altering one’s own surroundings and a move from the natural to the abstract in terms of design.
Summary of Continuing Modern H. sapiens in Africa
African culture experienced a long phase of development called the Middle Stone Age followed by a faster burst of change produced additional innovations and new styles. The change was not one moment but rather a ramping up in development. Later Stone Age culture introduced elements seen across many cultures, including the construction of composite tools and even the use of strung decorations such as beads. These developments appear in the Later Stone Age of other regions, such as with the Balangoda of Sri Lanka and the Aurignacian tradition of Europe. Based on the early date of the African artifacts, Later Stone Age culture likely originated in Africa and passed from person to person and region to region, with people adapting the general technique to their local resources and viewing the meaning in their own way. Unfortunately, information about modern humans in Africa from 40,000 to 12,000 years ago is scarce.
Discovering the Americas
By 20,000 years ago, our species was the only member of Homo left on Earth. Gone were the Neanderthals, Denisovans, and Homo floresiensis. The range of modern Homo sapiens kept expanding eastward into—using the name given to this area by Europeans much later—the Western Hemisphere. This section will address what we know about the peopling of the Americas, from the first entry to these continents to the rapid spread of prehistoric Native Americans across its lush and varied environments.
The Changing Role of Beringia
Evidence points to a prehistoric land bridge called Beringia that allowed people to cross from Asia to North America, just as expansion to Australia was made easier with lowered sea levels that exposed Sunda and Sahul. Beringia connected what is now northeastern Siberia with Alaska. What people did to cross this land bridge is still being investigated. Currently there are two competing models for this event, called the Ice-Free Corridor model and the Coastal Route model, though the latter has been gaining intriguing evidence.
For most of the 20th century, the accepted theory was that prehistoric northeast Asians (East Asians and Siberians) first expanded across Beringia inland through an ice-free corridor between glaciers that opened into the western Great Plains of the United States, just east of the Rocky Mountains, around 13,000 years ago. While life up north in the cold environment would have been harsh, migrating birds and an emerging forest might have provided sustenance as generations expanded through this land. These residents would have used a stone tool style that developed into the common Clovis style found later in North America.
In recent decades, researchers accumulated evidence against the ice-free corridor as the original path the first Native Americans took. For example, some archaeological sites around the Americas date to a time before the corridor was open. While one site with a date that contradicts the model could be dismissed as an error at first, several more sites were found that brought more scrutiny to the Ice-Free Corridor model. The route between glaciers was available later, and was likely used at that time, but there was already a more accessible path between the hemispheres.
The reconstruction of past geography, climate, and ecology led to the formation of the Coastal Route model that explains how people reached the Americas through Beringia. The new focus is the southern edge of the land bridge instead of its center: About 16,000 years ago, members of our species expanded along the coastline from northeast Asia, east through Beringia, and south down the Pacific Coast of North America while the inland was still sealed off by ice. Reconstructing the geography and climate of Beringia, the coast would have been free of ice at least part of the year by 16,000 years ago, earlier than when the ice-free corridor was completely opened. Studies of past ecology found that the coastal route would have provided abundant plant and animal resources for most of the path. Besides migrating birds, many useful fish (e.g., salmon), shellfish, mammals (e.g., whales, seals, and otters), and plants (e.g., seaweed) would have been available on the coast.
Other lines of evidence are also compatible with the Coastal Migration model. One indirect archaeological connection between prehistoric Japan and the Americas is a certain style of stone tool. Coastal tanged or stemmed lithics, which are stone points with a thin projection at the base, have been found in both of these distant regions (Erlandson and Braje 2011). The similarity could indicate a cultural tie. Genetic analysis of Native American DNA also shows shared ancestry with northeast Asians, linking them biologically as well.
While many factors such as reconstructions of climate and ecology indirectly support the Coastal Migration model, researchers have also begun to find archaeological sites along the route. That effort is complicated by the warming trend since 18,000 years ago that reduced glaciers and raised the sea level, so that much of the prehistoric coast that would have been occupied by the first coastal migrants to North America is currently over 100 meters underwater.
South through the Americas
However the first modern Homo sapiens reached the Western Hemisphere, the spread through the Americas was rapid. Multiple migration waves crossed from North to South America (Posth et al. 2018). Our species took advantage of the lack of hominin competition and the bountiful resources both along the coasts and inland. The Americas had their own wide array of megafauna, which included woolly mammoths, mastodons, camels, horses, ground sloths, giant tortoises, and—a favorite of researchers—a two-meter-tall beaver. The reason we cannot find these amazing animals today may be that ancient humans hunted them all to extinction, though climate chance may also play a role. Resources gained from these fauna must have been an important part of survival for people over 12,000 years ago. Several sites are notable for what they add to our understanding of American prehistory, including interactions with megafauna and other elements of the environment.
Monte Verde is a landmark site that shows that the human population had expanded down the whole vertical stretch of the Americas to Chile by 14,600 years ago, only a few thousand years after humans first entered the Western Hemisphere from Alaska. The site has been excavated by archaeologist Tom D. Dillehay and his team (2015), revealing fragile material culture that is rarely preserved, including human footprints, animal hides, and wooden tools.
Named after the town in New Mexico, the Clovis stone tool style is the first example of a widespread culture across much of North America, between 13,400 and 12,700 years ago. Instead of a stem-shaped base, Clovis points were fluted with two small projections, one on each end of the base, facing away from the head (Figure 26). The stone points found at this site match those found as far as the Canadian border and northern Mexico, and from the west coast to the east coast of the United States. Fourteen Clovis sites also contained the remains of mammoths or mastodons, suggesting that hunting megafauna with these points was an important part of life for the Clovis people. Other Clovis sites show that other types of hunting and gathering were important to people’s subsistence, too. After the spread of the Clovis style, it diversified into several regional styles, keeping some of the Clovis form but also developing their own unique touches.

Summary of Modern H. sapiens in the Americas
Research in prehistoric Native American origins found some surprising details, refining older models. Genetically, the migration can be considered one long period of movement, with splits into regional populations. This finding matches the sudden appearance of the homegrown Clovis culture, its rapid expansion, and the radiation of descendant cultures in North America. A few thousand years after arrival into the hemisphere, people had already covered the Americas from north to south.
The peopling of the Americas also had a lot of common elements with the prior spread of humans across Africa, Europe, Asia, and Australia. In all of these expansions, people explored new lands that tested both the cultural and biological adaptations of the pioneers. Besides stone tool technology, the use of ochre as decoration was seen from South Africa to South America. The coasts and rivers were likely avenues in the movement of people, artifacts, and ideas, outlining the land masses while providing access to varied environments. The presence of megafauna aided human success, but this resource was eventually depleted in many parts of the world.
The Big Picture: The Assimilation Model
How do researchers make sense of all of these modern Homo sapiens discoveries that cover over 300,000 years of time and stretch across every continent except Antarctica? How was modern Homo sapiens related to archaic Homo sapiens? Over the past few decades, paleoanthropologists have engaged in spirited debates based on their interpretation of the data.
One competing model was called the Out of Africa model or Recent African Origin model. Supporters of this model saw evidence that modern Homo sapiens first evolved in Africa, then expanded into the other continents without interaction with the archaic Homo sapiens of Europe and Asia. Researchers on this side noted that the oldest modern Homo sapiens fossils were found in Africa, suggesting that that continent was the origin. Genetic analysis found the same conclusion.
The other model was called Multiregionalism or the Multiregional Continuity model. The view of the data by scientists on this side was that modern Homo sapiens evolved from the archaic humans in Africa, Europe, and Asia simultaneously. Gene flow would have kept the species cohesive across the great distance while producing local variation as well. The multiregionalist experts pointed to the sharing of traits from Homo erectus, through archaic Homo sapiens, and then to the modern humans in different parts of the world as supporting their model.
Eventually, researchers noticed that both the Out of Africa model and the Multiregionalism model had elements that were supported by data and elements that were not supported. Taking the supported parts of each model and combining them formed an explanation that was more complicated, but explained much more of the scientific evidence. The merging of two models to form a better one suits the name of the improved version: the Assimilation model.
The Assimilation model proposes that modern Homo sapiens evolved in Africa first and expanded out (from the Out of Africa model) but also interbred with the archaic Homo sapiens they encountered outside Africa (from the Multiregionalism model) (Figure 27). True multiregionalism occurred just within Africa as the species evolved from a web of interactions between varied groups. As the modern human population expanded from Africa, they assimilated the alleles of archaic humans they encountered through interbreeding. The Assimilation model is powerful since it explains why Africa has the oldest modern human fossils, why early modern humans found in Europe and Asia bear a resemblance to the regional archaics, and why traces of archaic DNA can be found in our genomes today (Smith et al. 2017).
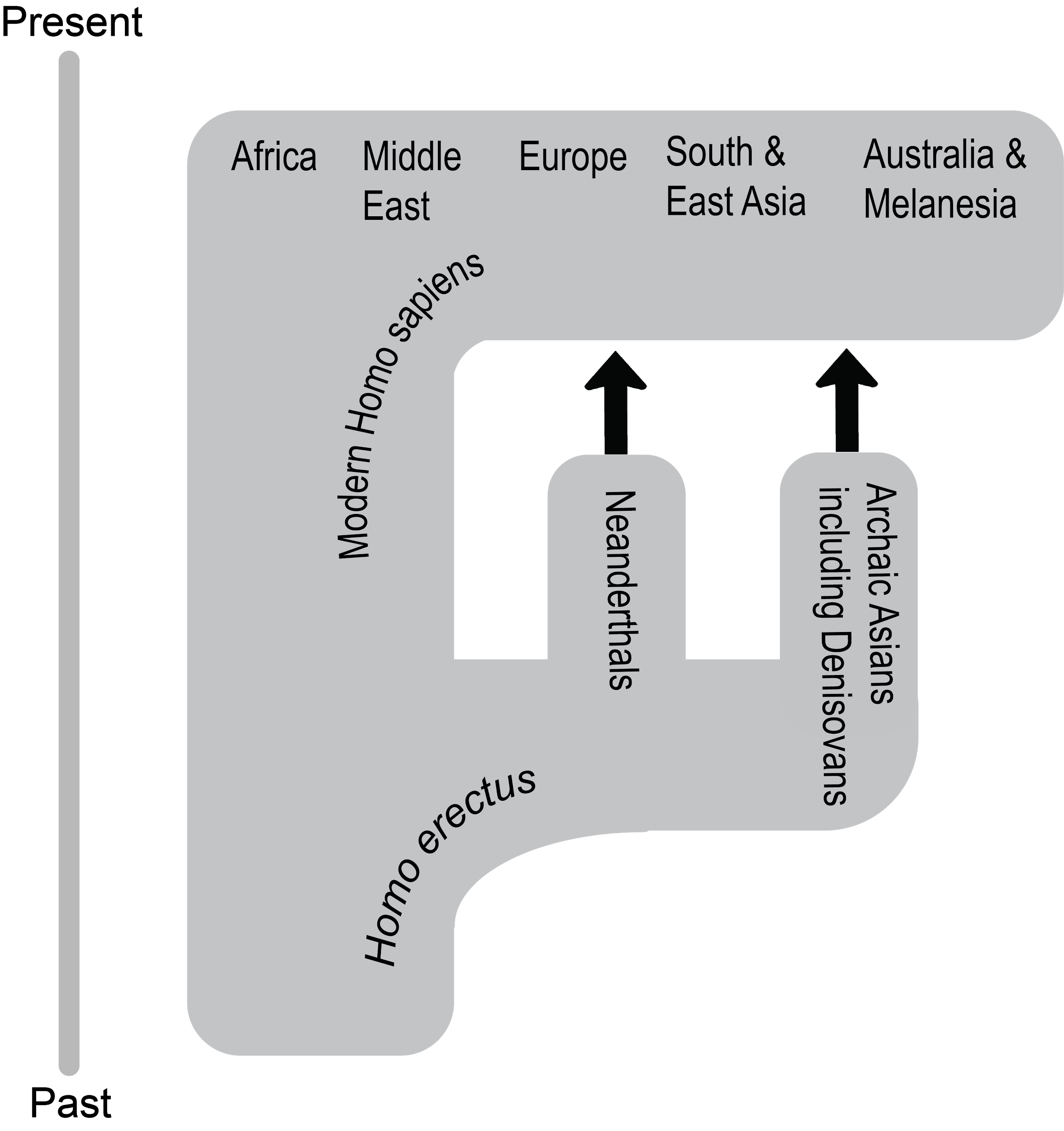
While scientific progress has produced a model that satisfies the data, there are still a lot of questions for paleoanthropologists to answer regarding our origins. What were the patterns of migration in each part of the world? Why did the archaic humans go extinct? In what ways did archaic and modern humans interact? How large were the past populations? How did biological, cultural, and environmental factors influence the material culture found in different parts of the world? The definitive explanation of how our species started and what our ancestors did is still out there to be found. You are now in a great place to welcome the next discovery about our distant past—maybe you’ll even contribute to our understanding as well.
“Cavemen” in Popular Culture
“Cavemen,” or our prehistory in general, is a constant presence in western popular culture. From the iconic opening to 2001: A Space Odyssey to the less iconic 10,000 B.C., the distant past is a common setting for dramatic stories. The prehistoric experience even gets interactive with games like Far Cry Primal where the player can persistence-hunt for food and ride a woolly mammoth (!) (Figure 28). The distant past has also been the setting for more comedic stories, such as The Flintstones and, recently, The Croods with Nicolas Cage.

Why are we interested in the past beyond a scientific understanding? Like feudal times or the Wild West, prehistory may be a setting that is rife for telling stories that excite us. With humanity stripped of the modern conveniences we are used to, situations become more intense. Prehistory takes this trope to the extreme. The viewer/game player is invited to think about what they would do if they had to live more directly off of the environment, unshielded from the dangers of the natural world. Experiencing our imaginings of that time may satisfy parts of our brain that evolved in that situation, just as people accustomed to urban life find enjoyment by camping or sitting around a fire with friends and family. Seeing the portrayal of the prehistoric world may reach those parts of our psychology that feel at home away from our constructed environment.
That said, the portrayal of the past in fiction is rarely accurate. There is a significant lag time between the scientific view of our past and when it reaches mass media. Older works age very poorly. In Clan of the Cave Bear, a book from 1980, the Neanderthals have limited emotions and communicate with sign language rather than speaking: two ideas that have not stood the test of time but were core to the story. Even in recent productions, prehistoric people are dim-witted and have limited vocabulary, though the evidence shows that our brains and language abilities are basically the same today as 300,000 years ago. How our media choose to represent the past may say more about our own values and views of ourselves rather than what people were actually like long ago.
As you engage in the next fiction set in our distant past, consider how the presentation differs from the scientific view and why there is this divide. Also, as you enjoy the world that is presented to you, consider that we are not so different from the cavemen. We are the ones born into the world of projectors and streaming media, and we may build on that to create the next medium, but at our core we are the same as the painters of Lascaux and the carvers of prehistoric figurines.
The Chain Reaction of Agriculture
While it may be hard to imagine today, for most of our species’ existence we were nomadic: moving through the landscape without a singular home. Instead of a refrigerator or pantry stocked with food, we procured nutrition and other resources as needed based on what was available in the environment. Instead of collecting and displaying stuff, we kept our possessions at a minimum for mobility. This foraging lifestyle enabled the expansion of our species, but the invention of a new way of life, caused a chain reaction of cultural change taking us to the present day and beyond.
The Foraging Tradition
To understand our species is to understand foraging, or the search for resources in the environment. This subsistence strategy, or method of finding sustenance, sounds unusual to most of us today. Most of us live in cultures that practice another strategy, agriculture, where we shape the environment to mass produce what we need. Considering the age of modern Homo sapiens, however, we have spent far more time as nomadic foragers than as settled agriculturalists. As such, our traits have evolved to be primarily geared toward foraging. For instance, our efficient bipedalism allows persistence-hunting across long distances as well as movement from resource to resource. Even our psychological tendency toward our ability to form stable relationships with around 150 people may derive from the foraging lifestyle.
How does human foraging, also known as hunting and gathering, work? Anthropologists have used all four fields to answer this question. Typically, people formed bands, or groups of around 50, and rarely over 100. A band’s organization would be egalitarian, with a flexible hierarchy based on an individual’s age, level of experience, and relationship with others. Everyone would have a general knowledge of the skills assigned to their gender roles, rather than specializing in different occupations. The ratio of plant to meat in one’s diet would have depended on the local resources. As a location’s resources became used up, and as human waste accumulated, the band would travel to another patch. In the varied environments that humans entered—from savannas to tropical forests, deserts, coasts, and the Arctic circle—people found sustenance needed for survival.
Why Agriculture?
After hundreds of thousands of years of foraging, some groups of people around 12,000 years ago started to practice agriculture instead. This transition is called the Neolithic Revolution, and it occurred at the start of the Holocene epoch. The reasons for this global change are still being investigated, but there are two likely causes that may have occurred together: a growing human population and natural global climate change.
Overcrowding could have affected the success of foraging in the environment, leading to the development of a more productive subsistence strategy. Foraging works the best with low population densities since each band needs a lot of space to support itself. If too many people occupy the same environment, they would deplete the area faster. This situation on a global level due to growing population and limited areas of expansion would have been an increasingly pressing issue after the human expansion through the major continents by 14,600 years ago.
A changing global climate immediately preceded the transition to agriculture, so researchers have also explored a connection between the two events. Since the Last Glacial Maximum of 23,000 years ago, the Earth slowly warmed. At 13,000 years ago, the temperature in most of the Northern Hemisphere dropped suddenly in a phenomenon called the Younger Dryas. Glaciers returned in Europe, Asia, and North America. In Mesopotamia, which includes the Levant, the climate changed from warm and humid to cool and dry. The change would have occurred over decades, disrupting the usual nomadic patterns and subsistence of foragers around the world. The disruption to foragers due to the temperature shift could have been a factor in spurring the transition to agriculture. Researchers Gregory K. Dow and colleagues (2009) believe that foraging bands would have clustered in the new resource-rich places where people started to direct their labor to farming the limited area. Continued practice would prompt innovations such as better tools and increasing productivity. After the Younger Dryas ended, people expanded out of the clusters. As they reinhabited the region, they brought with them a culture in which farming had become the norm, along with the technology and knowledge to succeed with this subsistence strategy.
The double threat of the limitation of human continental expansion and the sudden global climate change may have placed bands in peril as more populations outpaced their environment’s carrying capacity. Not only had a growing population led to increased competition with other bands, but environments worldwide shifted to create more uncertainty. As people in different areas around the world faced this chaotic situation, they became the independent inventors of agriculture.
Agriculture around the World
Due to global changes to the human experience starting from 12,000 years ago, cultures with no knowledge of each other turned toward farming their local resources. The switch to agriculture took time and effort with no guarantee of success. Agriculture is a difficult process with fires, floods, droughts, disease, and pests being constant problems to address. Heavy physical labor with no immediate payoff was also needed to shape the landscape in a coordinated way to support agriculture. For example, people had to direct water flow to irrigate constructed fields of crops. The first farmers also engaged in artificial selection of their domesticates to enhance useful traits. The biggest success stories in the face of these obstacles became the primary centers of agriculture:
- Mesopotamia: The Fertile Crescent from the Tigris and Euphrates rivers through the Levant was where bands started to domesticate plants and animals around 12,000 years ago. The connection between the development of agriculture and the Younger Dryas was especially strong here. Farmed crops included wheat, barley, peas, and lentils. This was also where cattle, pigs, sheep, and goats were domesticated.
- South and East Asia: Multiple regions across this land had varieties of rice, millet, and soybeans by 10,000 years ago. Pigs were farmed with no connection to Mesopotamia. Chickens were also originally from this region, bred for fighting first and food second.
- New Guinea: An under-appreciated center in Melanesia, agriculture started here 10,000 years ago. Bananas, sugarcane, and taro were native to this island. Sweet potatoes were brought back from voyages to South America around the year C.E. 1,000. No known animal farming occurred here.
- Mesoamerica: Agriculture from Central Mexico to northern South America also occurred from 10,000 years ago; it was also only plant based. Maize was a crop bred from teosinte grass, which has become one of the global staples. Beans, squash, and avocados were also grown in this region.
- The Andes: Starting around 8,000 years ago, local domesticated plants started with squash but later included potatoes, tomatoes, beans, and quinoa. Maize was brought down from Mesoamerica to join the local variety. The main farm animals were llamas, alpacas, and guinea pigs.
- Sub-Saharan Africa: This region went through a change 5,000 years ago called the Bantu expansion. The Bantu agriculturalists were established in West Central Africa and then expanded south and east. Native varieties of rice, yams, millet, and sorghum were grown across this area. Cattle were also domesticated here.
- Eastern North America: This region was the last major independent agriculture center, from 4,000 years ago. Squash and sunflower are the produce from this region that are most known today, though sumpweed and pitseed goosefoot were also farmed. Hunting was still the main source of animal products.
By 5,000 years ago, our species was well within the Neolithic Revolution. From the primary centers, agriculturalists spread to neighboring parts of the world with their domesticates, further expanding the use of this subsistence strategy. For example, the Mesopotamian farmers spread their innovations along the northern coast of the Mediterranean into Europe (Pinhasi, Fort, and Ammerman 2005). The crops of China were brought into the Korean peninsula (Lee 2011). From this point, the human species changed from being primarily foragers to primarily agriculturalists. The revolution took millennia, but it was a true revolution in that the lifestyle of our species was reshaped to something vastly different.
Conclusion
Modern Homo sapiens is the species that took the hominin lifestyle the furthest to become the only living member of that lineage. This last section of the chapter summarizes what we know about modern Homo sapiens traits, origins, and history.
The largest factor that allowed us to persist while other hominins went extinct was likely our advanced ability to culturally adapt to a wide variety of environments. Our species, with its skeletal and behavioral traits, was well suited to be generalist-specialists who successfully foraged across most of the world’s environments. The biological basis of this adaptation was our reorganized brain that facilitated innovation in cultural adaptations and intelligence for leveraging our social ties. As the brain’s ability increased, it shaped the skull by reducing the evolutionary pressure to have large teeth and robust cranial bones to produce the modern Homo sapiens face.
Our ability to be generalist-specialists is seen in the geographical range that modern Homo sapiens covered in 300,000 years. In Africa, our species formed from multiregional gene flow that loosely connected archaic humans across the continent. People then expanded out to the rest of the continental Old World and even further to the Americas. Wherever people went, they were enabled and connected by the shared tools and art they crafted.
For most of our species’ existence, foraging was the general subsistence strategy within which people specialized to culturally adapt to their local environment. With biologically endowed omnivorousness and mobility, people found ways to extract and process resources, shaping the environment in return. When global fluctuation in climate and a sudden resource uncertainty hit the species, people around the world focused on agriculture to have a firmer control of necessities. The new strategy shifted human history toward exponential growth and innovation to address the drastic shift in lifestyle, leading to our high dependence on cultural adaptations today. We may continue this trend in the future, with global changes to human genetic diversity.
While a cohesive image of our species has formed in recent years, there is still much to learn about our past. The work of many driven researchers shows that there are amazing new discoveries made all the time that refine our knowledge of human evolution. Technological innovations such as DNA analysis enable scientists to approach lingering questions from new angles. The answers we get allow us to ask even more insightful questions that will lead us to the next revelation. Like the pink limestone strata at Jebel Irhoud, previous effort has taken us so far and you are now ready to see what the next layer of discovery holds.
Terms You Should Know
Study Questions
- What are the features used to define modern humans?
- What are the most likely scenarios for how modern humans evolved?
- How do the discoveries mentioned in “First Africa, Then the World” fit the Assimilation model?
- What is foraging? What adaptations do we have for this subsistence strategy? Could you train to be a skilled forager?
References
Berger, Thomas D., and Erik Trinkaus. 1995. Patterns of Trauma among the Neanderthals. Journal of Archaeological Science 22(6): 841–852.
Bocherens, Hervé, Daniel Billiou, André Mariotti, Marylène Patou-Mathis, Marcel Otte, Dominique Bonjean, and Michel Toussaint. 1999. Palaeoenvionmental and Palaeodietary Implications of Isotopic Biogeochemistry of Last Interglacial Neanderthal and Mammal Bones in Scladina Cave (Belgium). Journal of Archaeological Science 26(6): 599–607.
Bowler, J. M., Rhys Jones, Harry Allen, and A. G. Thorne. 1970. Pleistocene Human Remains from Australia: A Living Site and Human Cremation from Lake Mungo, Western New South Wales. World Archaeology 2 (1): 39–60.
Dannemann, Michael, and Fernando Racimo. 2018. Something Old, Something Borrowed: Admixture and Adaptation in Human Evolution. Current Opinion in Genetics & Development 53: 1–8.
Defleur, Alban R., and Emmanuel Desclaux. 2019. Impact of the Last Interglacial Climate Change on Ecosystems and Neanderthals Behavior at Baume Moula-Guercy, Ardèche, France. Journal of Archaeological Science 104: 114–124.
Degioanni Anna, Chritophe Bonenfant, Sandrine Cabut, and Silvana Condemi. 2019. Living on the Edge: Was Demographic Weakness the Cause of Neanderthal Demise? PLoS ONE 14 (5): e0216742.
Dillehay, Tom D., Carlos Ocampo, José Saavedra, Andre Oliveira Sawakuchi, Rodrigo M. Vega, Mario Pino, Michael B. Collins, et al. 2015. New Archaeological Evidence for an Early Human Presence at Monte Verde, Chile. PLOS ONE 10(11): e0141923.
Dow, Gregory K., Clyde G. Reed, and Nancy Olewiler. 2009. Climate Reversals and the Transition to Agriculture. Journal of Economic Growth 14(1): 27–53.
Erlandson, Jon M., Todd J. Braje, Kristina M. Gill, and Michael H. Graham. 2015. Ecology of the Kelp Highway: Did Marine Resources Facilitate Human Dispersal from Northeast Asia to the Americas? The Journal of Island and Coastal Archaeology 10(3): 392–411.
Golovanova, Liubov. Vitaliena, Vladimir B. Doronichev, Naomi E. Cleghorn, Marianna A. Koulkova, Tatiana V. Sapelko, and M. Steven Shackley. 2010. Significance of Ecological Factors in the Middle to Upper Paleolithic Transition. Current Anthropology 51(5): 655–691.
Henshilwood, Christopher S., Francesco d’Errico, Karen L. van Niekerk, Yvan Coquinot, Zenobia Jacobs, Stein-Erik Lauritzen, Michel Menu, and Renata García-Moreno. 2011. A 100,000-Year-Old Ochre-Processing Workshop at Blombos Cave, South Africa. Science 334(6053): 219–222.
Houldcroft, Charlotte J., and Simon J. Underdown. 2016. Neanderthal Genomics Suggests a Pleistocene Time Frame for the First Epidemiologic Transition. American Journal of Physical Anthropology 160 (3): 379–388.
Hublin, Jean-Jacques, Abdelouahed Ben-Ncer, Shara E. Bailey, Sarah E. Freidline, Simon Neubauer, Matthew M. Skinner, Inga Bergmann, et al. 2017. New Fossils from Jebel Irhoud, Morocco, and the Pan-African Origin of Homo sapiens. Nature 546(7657): 289–292.
Kortlandt, Adriaan. 2002. Neanderthal Anatomy and the Use of Spears. Evolutionary Anthropology 11(5): 183–184.
Lieberman, Daniel E., Brandeis M. McBratney, and Gail Krovitz. 2002. The Evolution and Development of Cranial Form in Homo sapiens. Proceedings of the National Academy of Sciences 99(3): 1134–1139.
Liu, Wu, María Martinón-Torres, Yan-jun Cai, Song Xing, Hao-wen Tong, Shu-wen Pei, Mark Jan Sier, Xiao-hong Wu, R. Lawrence Edwards, and Hai Cheng. 2015. The Earliest Unequivocally Modern Humans in Southern China. Nature 526(7575): 696-699.
McDougall, Ian, Francis H. Brown, and John G. Fleagle. 2005. Stratigraphic Placement and Age of Modern Humans from Kibish, Ethiopia. Nature 433(7027): 733–736.
Neubauer, Simon, Jean-Jacques Hublin, and Philipp Gunz. 2018. The Evolution of Modern Human Brain Shape. Science Advances 4 (1): eaao5961.
Pearce, Eiluned, Chris Stringer, and R. I. M. Dunbar. 2013. New Insights into Differences in Brain Organizations between Neanderthals and Anatomically Modern Humans. Proceedings of the Royal Society B: Biological Sciences 280(1758): 20130168.
Pietschnig, Jakob, Lars Penke, Jelte M. Wicherts, Michael Zeiler, and Martin Voracek. 2015. Meta-Analysis of Associations between Human Brain Volume and Intelligence Differences: How Strong Are They and What Do They Mean? Neuroscience & Biobehavioral Reviews 57: 411–432.
Posth, Cosimo, Nathan Nakatsuka, Iosif Lazaridis, Pontus Skoglund, Swapan Mallick, Thiseas C. Lamnidis, Nadin Rohland, et al. 2018. Reconstructing the Deep Population History of Central and South America. Cell 175(5): 1185–1197.
Scerri, Eleanor M. L., Mark G. Thomas, Andrea Manica, Philipp Gunz, Jay T. Stock, Chris Stringer, Matt Grove, et al. 2018. Did Our Species Evolve in Subdivided Populations Across Africa, and Why Does It Matter? Trends in Ecology & Evolution 33(8): 582–594.
Shea, John J. 2011. Refuting a Myth about Human Origins. American Scientist 99(2): 128–135.
Shea, John J., and Ofer Bar-Yosef. 2005. Who Were the Skhul/Qafzeh People? An Archaeological Perspective on Eurasia’s Oldest Modern Humans. Journal of the Israel Prehistoric Society 35: 451–468.
Smith, Fred H., James C. M. Ahern, Ivor Janković, and Ivor Karavanić. 2017. The Assimilation Model of Modern Human Origins in Light of Current Genetic and Genomic Knowledge. Quaternary International 450: 126–136.
Staubwasser, Michael, Virgil Drăguʂin, Bogdan P. Onac, Sergey Assonov, Vasile Ersek, Dirk L. Hoffman, and Daniel Veres. 2018. Impact of Climate Change on the Transition of Neanderthals to Modern Humans in Europe. Proceedings of the National Academy of Sciences 115(37): 9116–9121.
Trinkaus, Erik, Ştefan Milota, Ricardo Rodrigo, Gherase Mircea, and Oana Moldovan. 2003. Early Modern Human Cranial Remains from the Peştera Cu Oase, Romania. Journal of Human Evolution 45(3): 245–253.
Wall, Jeffrey D., and Deborah Yoshihara Caldeira Brandt. 2016. Archaic Admixture in Human History. Current Opinion in Genetics & Development 41: 93–97.
Weaver, T. D., and J. Hublin. 2009. “Neanderthal Birth Canal Shape and the Evolution of Human Childbirth.” Proceedings of the National Academy of Sciences 106 (20): 8151–8156.
White, Tim D., Berhane Asfaw, David DeGusta, Henry Gilbert, Gary D. Richards, Gen Suwa, and F. Clark Howell. 2003. Pleistocene Homo sapiens from Middle Awash, Ethiopia. Nature 423(6941): 742–747.
A Derivative Work From
Chan, Keith. 2023. Modern Homo Sapiens. In Shook, Beth; Nelson, Katie; Aguilera, Kelsie; and Braff, Lara (eds.). Explorations: An Open Invitation to Biological Anthropology, Second Edition. American Anthropological Association.
Wolcott Paskey, Amanda and Beasley Cisneros, AnnMarie. 2023. Archaic Homo. In Shook, Beth; Nelson, Katie; Aguilera, Kelsie; and Braff, Lara (eds.). Explorations: An Open Invitation to Biological Anthropology, Second Edition. American Anthropological Association.
Having a rounded appearance. Increased globularity of the braincase is a trait of modern Homo sapiens.
Australopithecine species with rugged or exaggerated features, especially related to teeth and the masticatory system (i.e. chewing).
Stone tools made and used by Neanderthals. These tools were made using the prepared-core technique, were of high quality, and were saved and reused.
A natural clay pigment mixed with ferric oxide and clay and sand. Ranges in color from brown to red to orange. Found in Neanderthal burials.
A form of dwarfism that occurs when a limited geographic region, such as an island, causes a large-bodied animal to be selected for a smaller body size.
Composed from a mix or composite of traits.
Having a smooth and slender quality; the opposite of robust.
In reference to the face, the area below the eyes juts anteriorly.
Time period known for Mousterian-style lithics, and which connects African archaic to modern Homo sapiens temporally.
Time period following the Middle Stone Age with a diversification in tool types, starting around 50,000 years ago.
The idea that modern Homo sapiens evolved as a complex web of small regional populations in Africa, with sporadic gene flow among them.
Large ancient animals that may have been hunted to extinction by people around the world.
The uneven mixing of DNA over time in which a small amount of outside genetic material is incorporated into a larger genome.
Small stone tool found in the Later Stone Age; also called a bladelet.
Prehistoric landmass that connected Siberia and Alaska. The ancestors of Native Americans would have crossed this area to reach the Americas.
Theory that the first prehistoric Native Americans crossed to the Americas through a passage between glaciers.
Theory that the first Paleoindians crossed to the Americas by following the southern coast of Beringia.
Theory that modern Homo sapiens expanded from Africa to cover the rest of the world by replacing and without interacting with archaic humans.
Theory that modern Homo sapiens evolved simultaneously in Africa, Asia, and Europe from archaic populations.
Current theory of modern human origins stating that the species evolved first in Africa and interbred with archaic humans of Europe and Asia.
Describes populations that move through the landscape without a singular home.
a mode of subsistence defined by its reliance on wild plant and animal food resources already available in the environment rather than on domesticated species that have been altered by human intervention.
the set of skills, practices, and technologies used by members of a society to acquire and distribute food.
The cultivation of domesticated plants and animals using technologies such as irrigation, draft animals, mechanization, and inputs such as fertilizers and pesticides that allow for intensive and continuous use of land resources.
the smallest unit of political organization, consisting of only a few families and no formal leadership positions.
societies in which there is no great difference in status or power between individuals and there are as many valued status positions in the societies as there are persons able to fill them.
a period of rapid innovation in subsistence technologies that began 10,000 years ago and led to the emergence of agriculture. Neolithic means “new stone age,” a name referring to the stone tools produced during this time period.
The epoch of the Cenozoic Era starting around 12,000 years ago and lasting arguably through the present.
The rapid change in global climate, especially a cooling of the Northern Hemisphere, 13,000 years ago.
