
16
Learning Goals
By the end of this reading you should be able to:
- Explain the advantages of simple multicellularity
- Describe the evidence that supports the evolution of simple multicellularity in some protists
- Discuss the needed adaptations for simple multicellularity
Introduction
Among the Bacteria and Archaea, each cell usually functions as an individual, growing and reproducing, moving from one place to another, taking in nutrients, and both sensing and transmitting molecular signals. In contrast, multicellular organisms contain millions (or more) cells that work in close coordination. In your own body, for example, different cells are specialized for specific functions, so that while your body as a whole can perform the broad range of tasks accomplished by microorganisms, individual cells, for the most part, cannot. Cells lining the intestine absorb food molecules, but nutrients must then be transported to other parts of the body. Lungs take up oxygen from inhaled air, but this, too, must be distributed to other tissues and organs. And while cells at the body’s surface sense signals from the environment, the signals affecting interior cells come mostly from surrounding cells. While the biological gulf between microbes and multicellular organisms is enormous, multicellularity has evolved a half dozen times in different groups.
The phylogeny of multicellularity
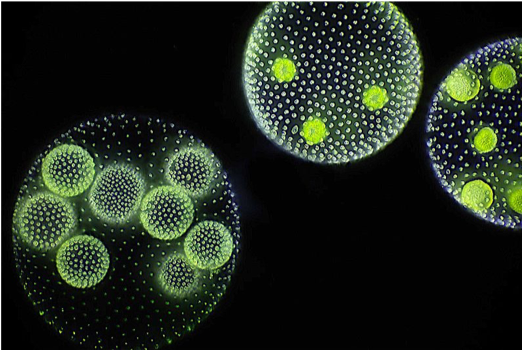
Most prokaryotic organisms are composed of a single cell, although some form simple filaments or live in colonies. A few types of bacterium, notably some cyanobacteria, differentiate to form several distinct cell types. No bacteria, however, develop macroscopic bodies with functionally differentiated tissues. Only eukaryotes have evolved those. Simple multicellular organisms, composed of multiple similar cells, occur widely within the eukaryotic tree of life, and a few of these evolved into more complex multicellular organisms characterized by differentiated tissues and organs. How are these different types of organization distribute on the eukaryotic tree of life?
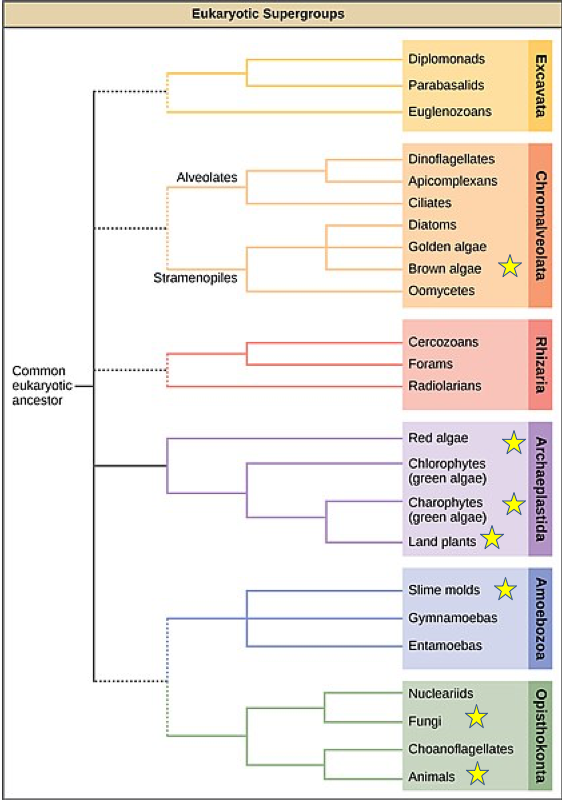
A recent survey of eukaryotic organisms recognized 119 major groups within the supergroups. Of these, 83 contain only single-celled organisms, predominantly cells that engulf other microorganisms or ingest small organic particles, photosynthetic cells that live suspended in the water column, or parasitic cells that live within other organisms. Each of the 36 remaining branches exhibits some cases of simple multicellularity, mostly in the form of filaments, hollow balls, or sheets of little-differentiated cells. More complex multicellularity has evolved only in six eukaryotic groups: animals, fungi, brown algae, red algae, green algae, and land plants.
Thinking Question:
Advantages of simple multicellularity
What selection did pressures favor the evolution of simple multicellular organisms from single-celled ancestors? One selective advantage is that multicellularity helps organisms avoid getting eaten. A number of experiments using single-celled green algae and exposure to a single cell predator have demonstrated the selection pressure for multicellularity. In the presence of the predators, within 10 to 20 generations, most of the algae were living in eight-cell colonies that were essentially invulnerable to predator attacks due to their larger size.1,2
Another advantage is that multicellular organisms may be able to maintain their position on a surface or in the water column better than their single-celled relatives. Seaweeds, for example, live anchored to the seafloor in places where light and nutrients support growth. Scientists have studied these phenomena using both single-celled algae and yeast cultures.3,4 When exposed to consistent shaking in culture, the number of cells that formed aggregates (clumps) increases over a number of generations. By clumping together the cell groups settled on the bottom of the cultures and thus were exposed to less vigorous conditions.
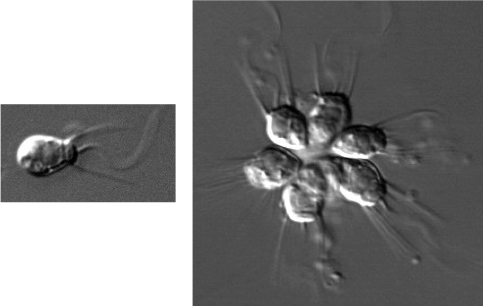
Feeding provides a third potential advantage: In colonial heterotrophs such as the stalked ciliate Epistylis, the coordinated beating of flagella assists feeding by directing currents of food-laden water toward the cells. Studies using the choanoflagellate Salpingoeca rosetta, demonstrated a link between the presence of the prey species and the development of multicellular colonies (formation of a rosette). These colonies, unlike the ones in the yeast and algae mentioned earlier, appear to the result of cell division rather than cell aggregation.5
Adaptations need for multicellularity
The genome of the choanoflagellate Monosiga brevicollis provides a fascinating insight into the evolution of cell adhesion molecules. Choanoflagellates, the closest protistan relatives of animals, are unicellular microorganisms. Therefore, it came as a surprise that the genes of M. brevicollis code for many of the same protein families that promote cell adhesion in animals. This includes the genes for both cadherin and integrin proteins which in animals are involved in supporting epithelia. Clearly, these proteins are not supporting epithelia in M. brevicollis, so what are they doing?
One approach to an answer came from the observation that cells stick not only to one another but also to rock or sediment surfaces. Proteins that originally evolved to promote adhesion of individual cells to sand grains or a rock surface may have been modified during evolution for cell-cell adhesion. It is also possible that adhesion molecules were originally used in the capture of bacterial cells, to make prey adhere to the predator.
To date, cadherins have been found only in choanoflagellates and animals, but proteins of the integrin complex extend even deeper into eukaryotic phylogeny-they are found in single celled protists that branch near the base of the opisthokont superkingdom. Such a phylogenetic distribution provides strong support for the general hypothesis that cell adhesion in animals resulted from the redeployment of protein families that evolved to perform other functions before animals diverged from their closest protistan relatives.
Thinking Question:
Adhesion but no differentiation
In many protists, cell adhesion molecules cause adjacent cells to stick together, but there are few specialized cell types and relatively little communication or transfer of resources between cells. Most or all of the cells in simple multicellular organisms retain a full range of functions, including reproduction. As a result the organism/colony usually pays only a small penalty for individual cell death. Importantly, in simple multicellular organisms, nearly every cell is in direct contact with the external environment, at least during phases of the life cycle when the cells must acquire nutrients, which means that each can take up nutrients and excrete wastes directly.
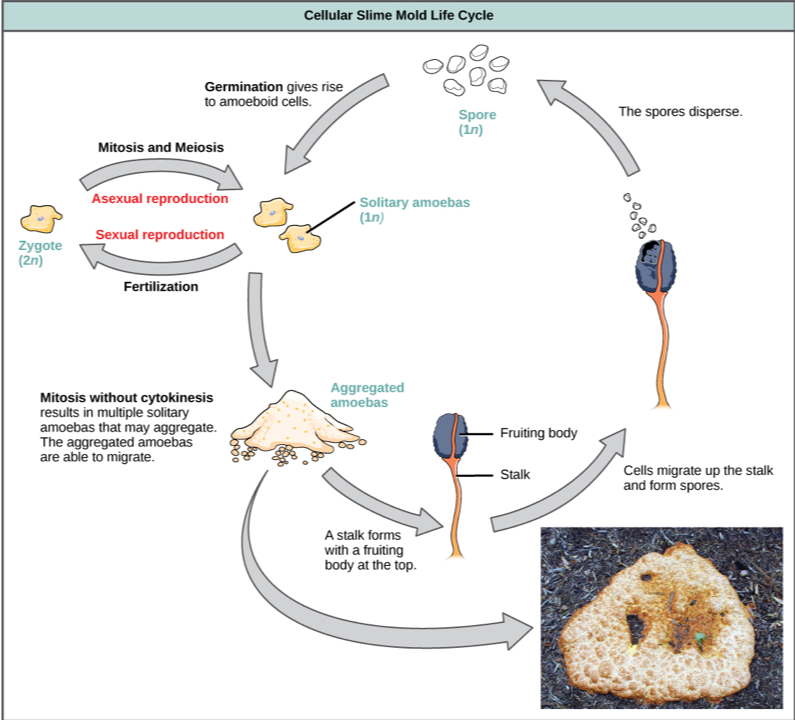
Simple multicellularity occurs most prominently among algae, although stalk-like colonies of particle feeders have evolved in at least three groups of heterotrophic protists. In addition, simple filamentous fungi exist that can absorb organic molecules as sources of carbon and energy. Four eukaryotic groups have achieved temporary simple multicellularity by a different route, aggregating during just one stage of the life cycle. Slime molds are the best-known example.
Six groups (two algal, three protozoan, and one fungal) include species characterized by coenocytic organization. In coenocytic organisms, the nucleus divides multiple times, but the cell does not, so the nuclei are not partitioned into individual cells. The result is a large cell-sometimes even visible to the naked eye with many nuclei. The green algae Codium and Caulerpa, found along the shorelines of temperate and tropical seas, respectively, are common examples of coenocytic organisms. In addition, acellular slime molds are also coenocytic and can be so large that they look like fungi to us whereas they are only one really really large cell. There is no evidence that any coenocytic organisms evolved from truly multicellular ancestors, nor have any given rise to complex multicellular descendants.
Review Question:
Summary
Given the benefits associated with being multicellular, it is no surprise that multiple lineages independently evolved multicellularity over time. This likely took advantage of traits that were already present in the organisms to help those ancestral cells adhere to a substrate. The first multicellular organisms were likely simple and the cells merely stuck together but did not differentiate and have different functions for the whole. Other lineages evolved processes in which the nucleus divides multiple times in the same cell, forming a coenocytic cell, but there is no evidence that multicellularity evolved from coenocytic cells.
End of Section Review Questions:
2) What do features would you expect SIMPLE multicellular organisms have in common?
References
1 Boraas, M. E., D. B. Seale, and J. E. Boxhorn. 1998. Phagotrophy by a flagellate selects for colonial prey: a possible origin of multicellularity. Evol. Ecol. 12:153–164 https://link.springer.com/article/10.1023/A:1006527528063
2 Matthew D. Herron, Josh M. Borin, Jacob C. Boswell, Jillian Walker, I-Chen Kimberly Chen, Charles A. Knox, Margrethe Boyd, Frank Rosenzweig, William C. Ratcliff. 2018. De novo origin of multicellularity in response to predation bioRxiv 247361; doi: https://doi.org/10.1101/247361
3 Ratcliff, W. C., Denison, R. F., Borrello, M. & Travisano, M. Proc. Natl Acad. Sci. USA advance online publication http://dx.doi.org/10.1073/pnas.1115323109 (2012).
4 Ratcliff, William C., et al. “Experimental evolution of an alternating uni-and multicellular life cycle in Chlamydomonas reinhardtii.” Nature communications 4 (2013): 2742. https://www.nature.com/articles/ncomms3742
5 Fairclough SR, Dayel MJ, King N. Multicellular development in a choanoflagellate. Curr Biol. 2010;20(20):R875-6. doi: 10.1016/j.cub.2010.09.014
Image attribution
Figure 1. Image courtesy of Frank Fox [CC BY-SA 3.0 de (https://creativecommons.org/licenses/by-sa/3.0/de/deed.en)], via Wikimedia Commons
Figure 2. Image courtesy of CNX OpenStax [CC BY 4.0 (https://creativecommons.org/licenses/by/4.0)], via Wikimedia Commons
Figure 3. Image courtesy of Mark J. Dayel [CC BY-SA 3.0 (https://creativecommons.org/licenses/by-sa/3.0)], via Wikimedia Commons
Figure 4. Image courtesy of CNX OpenStax [CC BY 4.0 (https://creativecommons.org/licenses/by/4.0)], via Wikimedia Commons
")
A YouTube element has been excluded from this version of the text. You can view it online here: https://viva.pressbooks.pub/introbio2/?p=27
