
3 Evolution

All organisms are products of evolution adapted to their environment. On the left, Saguaro (Carnegiea gigantea) can soak up 750 liters of water in a single rain storm, enabling these cacti to survive the dry conditions of the Sonora desert in Mexico and the Southwestern United States. On the right, the Andean semiaquatic lizard (Potamites montanicola) discovered in Peru in 2010 lives between 1,570 to 2,100 meters in elevation, and, unlike most lizards, is nocturnal and swims. Scientists still do not know how these cold-blood animals are able to move in the cold (10 to 15°C) temperatures of the Andean night. (credit a: modification of work by Gentry George, U.S. Fish and Wildlife Service; credit b: modification of work by Germán Chávez and Diego Vásquez, ZooKeys)
The theory of evolution is the unifying theory of biology, meaning it is the framework within which biologists ask questions about the living world. Because ecology is a branch of biology, understanding evolution is crucial for understanding ecology. In Chapter 1, we talked about scientific theories and described some theories as fundamental theories. Evolution is one of these fundamental theories because of its power to explain a lot about our world and its support from repeated observations and experiments. Its power is that it provides direction for predictions about living things that are borne out in ongoing experiments. The Ukrainian-born American geneticist Theodosius Dobzhansky famously wrote that “nothing makes sense in biology except in the light of evolution.”1 He meant that the tenet that all life has evolved and diversified from a common ancestor is the foundation from which we approach all questions in biology. Evolution is a change in the inherited characteristics (what we now know is the genetic makeup) of a species through generations. Many people confuse natural selection (explained below) and human evolution with the overall theory of evolution. These two concepts are certainly part of our understanding of evolution, but do not encompass all of the theory of evolution.
Development of the Theory of Evolution
Evolution by natural selection describes a mechanism for how species change over time. Scientists, philosophers, researchers, and others had made suggestions and debated this topic well before Darwin began to explore this idea. Classical Greek philosopher Plato emphasized in his writings that species were static and unchanging, yet there were also ancient Greeks who expressed evolutionary ideas. In the eighteenth century, naturalist Georges-Louis Leclerc Comte de Buffon reintroduced ideas about the evolution of animals and observed that various geographic regions have different plant and animal populations, even when the environments are similar. Some at this time also accepted that there were extinct species.
Also during the eighteenth century, James Hutton, a Scottish geologist and naturalist, proposed that geological change occurred gradually by accumulating small changes from processes operating like they are today over long periods of time, a theory called uniformitarianism. This contrasted with the predominant view that the planet’s geology was a consequence of catastrophic events occurring during a relatively brief past, a theory called catastrophism. For example, from a uniformitarian viewpoint, the Grand Canyon was formed over millions of years from the slow erosion of sediment by the Colorado River. From the catastrophism perspective, the Grand Canyon formed from a single large event, unlike events we see today. Nineteenth century geologist Charles Lyell popularized Hutton’s view, which is accepted today. A friend to Darwin. Lyell’s ideas were influential on Darwin’s thinking: Lyell’s notion of the greater age of Earth gave more time for gradual change in species, and the process of change provided an analogy for this change. In the early nineteenth century, Jean-Baptiste Lamarck published a book that detailed a mechanism for evolutionary change. Lamarck proposed that changes in an individual organism during its lifetime were passed on to its offspring in a theory of inheritance of acquired characteristics. The classic example from Lamarck’s theory is that giraffes repeatedly stretched their necks to reach higher leaves on trees. As the giraffe stretched its neck, its neck got longer and this longer neck was passed on to future generations. While Lamarck’s theory was discredited, in recent years, the study of epigenetics has demonstrated that behaviors and the environment of an individual can affect the expression of that individual’s DNA with the possibility that these changes can be passed on through generations. You can read more at this website with information about epigenetics.
Charles Darwin and Natural Selection
In the mid-nineteenth century, two naturalists, Charles Darwin and Alfred Russel Wallace, independently conceived and described natural selection as a mechanism for evolution. Importantly, each naturalist spent time exploring the natural world on expeditions to the tropics. From 1831 to 1836, Darwin traveled around the world on H.M.S. Beagle, including stops in South America, Australia, and the southern tip of Africa. Wallace traveled to Brazil to collect insects in the Amazon rainforest from 1848 to 1852 and to the Malay Archipelago from 1854 to 1862. Darwin’s journey, like Wallace’s later journeys to the Malay Archipelago, included stops at several island chains, the last being the Galápagos Islands west of Ecuador. On these islands, Darwin observed species of organisms on different islands that were clearly similar, yet had distinct differences. For example, the ground finches inhabiting the Galápagos Islands comprised several species with a unique beak shape. The species on the islands had a graded series of beak sizes and shapes with very small differences between the most similar. He observed that these finches closely resembled another finch species on the South American mainland. Darwin imagined that the island species might be species modified from one of the original mainland species. Upon further study, he realized that each finch’s varied beaks helped the birds acquire a specific type of food. For example, seed-eating finches had stronger, thicker beaks for breaking seeds, and insect-eating finches had spear-like beaks for stabbing their prey.
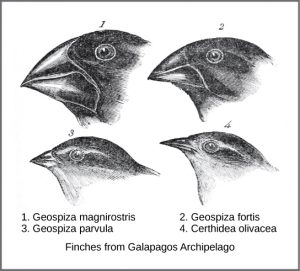 Darwin observed that beak shape varies among finch species. He postulated that ancestral species’ beaks had adapted over time to equip the finches to acquire different food sources.
Darwin observed that beak shape varies among finch species. He postulated that ancestral species’ beaks had adapted over time to equip the finches to acquire different food sources.
Wallace and Darwin both observed similar patterns in other organisms and they independently developed the same explanation for how and why such changes could take place. Darwin called this mechanism natural selection. Natural selection is the more prolific reproduction of individuals with favorable traits that survive environmental change because of those traits. This leads to evolutionary change. Natural selection is sometimes referred to as “survival of the fittest,” but this phrase does not accurately represent natural selection. Natural selection requires that an individual not only survive, but also reproduce and that the characteristics that increased the likelihood of that individual surviving and reproducing are passed on to the next generation.
For example, Darwin observed a population of giant tortoises in the Galápagos Archipelago to have longer necks than those that lived on other islands with dry lowlands. These tortoises were “selected” because they could reach more leaves and access more food than those with short necks. In times of drought when fewer leaves would be available, those that could reach more leaves had a better chance to eat and survive than those that couldn’t reach the food source. Consequently, long-necked tortoises would be more likely to be reproductively successful and pass the long-necked trait to their offspring. Over time, only long-necked tortoises would be present in the population.
Natural selection, Darwin argued, was an inevitable outcome of three principles that operated in nature. First, most characteristics of organisms are inherited, or passed from parent to offspring. Although no one, including Darwin and Wallace, knew how this happened at the time, it was a common understanding. Second, more offspring are produced than are able to survive, so resources for survival and reproduction are limited. The capacity for reproduction in all organisms outstrips the availability of resources to support their numbers. Thus, there is competition for those resources in each generation. Both Darwin and Wallace’s understanding of this principle came from reading economist Thomas Malthus’ essay that explained this principle in relation to human populations. Third, offspring vary among each other in regard to their characteristics and those variations are inherited. Darwin and Wallace reasoned that offspring with inherited characteristics which allow them to best compete for limited resources will survive and have more offspring than those individuals with variations that are less able to compete. Because characteristics are inherited, these traits will be better represented in the next generation. This will lead to change in populations over generations in a process that Darwin called descent with modification. Ultimately, natural selection leads to greater adaptation of the population to its local environment. It is the only mechanism known for adaptive evolution.
In 1858, Darwin and Wallace (see picture at right) presented  papers at the Linnean Society in London that discussed the idea of natural selection. The following year Darwin’s book, On the Origin of Species, was published. His book outlined in considerable detail his arguments for evolution by natural selection.
papers at the Linnean Society in London that discussed the idea of natural selection. The following year Darwin’s book, On the Origin of Species, was published. His book outlined in considerable detail his arguments for evolution by natural selection.
It is difficult and time-consuming to document and present examples of evolution by natural selection. The Galápagos finches are an excellent example. Peter and Rosemary Grant and their colleagues have studied Galápagos finch populations every year since 1976 and have provided important evidence of natural selection. The Grants found changes from one generation to the next in beak shape distribution with the medium ground finch on the Galápagos island of Daphne Major. The birds have inherited a variation in their bill shape with some having wide deep bills and others having thinner bills. During a period in which rainfall was higher than normal because of an El Niño, there was a lack of large hard seeds that the large-billed birds ate; however, there was an abundance of the small soft seeds which the small-billed birds ate. Therefore, the small-billed birds were able to survive and reproduce. In the years following this El Niño, the Grants measured beak sizes in the population and found that the average bill size was smaller. Since bill size is an inherited trait, parents with smaller bills had more offspring and the bill evolved into a much smaller size. As conditions improved in 1987 and larger seeds became more available, the trend toward smaller average bill size ceased. The Grant’s were able to document evolution by natural selection on a much shorter time scale than many scientists have been able to demonstrate.
Processes and Patterns of Evolution
Natural selection can only take place if there is variation, or differences, among individuals in a population. Importantly, these differences must have some genetic basis; otherwise, the selection will not lead to change in the next generation. This is critical because nongenetic reasons can also cause variation among individuals such as an individual’s height because of better nutrition rather than different genes.
Before proceeding, if you need a quick review of basic genetics, take a break to watch this video about alleles and genes.
Genetic diversity in a population comes from two main mechanisms: mutation and sexual reproduction. Mutation, a change in DNA, is the ultimate source of new genetic variation in any population. The genetic changes that mutation causes can have one of three outcomes on the phenotype, the observable characteristics of the organism.
- 1) A mutation affects the organism’s phenotype in a way that gives it reduced fitness—lower likelihood of survival or fewer offspring.
- 2) A mutation may produce a phenotype with a beneficial effect on fitness.
- 3) Many mutations, called neutral mutations, will also have no effect on the phenotype’s fitness.
Mutations also vary in how large their impact is on the organism’s fitness that expresses them in their phenotype, from a small effect to a great effect. Sexual reproduction also leads to genetic diversity: when two parents reproduce, unique combinations of genetic material assemble to produce the unique characteristics of the offspring. Sexual reproduction, such as pollination of flowers and fertilization of eggs in animals, involves the combination of genetic material from two organisms. In asexual reproduction, a single organism produces offspring, such as the runners from a strawberry plant producing new plants.
We call a heritable trait that helps an organism’s survival and reproduction in its present environment an adaptation. Scientists describe groups of organisms adapting to their environment when a genetic variation occurs over time that increases or maintains the population’s “fit” to its environment. A platypus’s webbed feet are an adaptation for swimming. A snow leopard’s thick fur is an adaptation for living in the cold. A cheetah’s fast speed is an adaptation for catching prey.
Whether or not a trait is favorable depends on the current environmental conditions. The same traits are not always selected because environmental conditions can change. For example, consider a plant species that grew in a moist climate and did not need to conserve water. Large leaves were selected because they allowed the plant to obtain more energy from the sun and produce more offspring in the next generation, therefore increasing the overall leaf size of the species. Large leaves require more water to maintain than small leaves, and the moist environment provided favorable conditions to support large leaves. After thousands of years, the climate changed, and the area no longer had excess water. The direction of natural selection shifted so that plants with small leaves were selected because those populations were able to conserve water to survive the new environmental conditions. If the plant does not adapt, the plant may go extinct in that area. A plant, or any organism, cannot choose to adapt because of changing environmental conditions, adaptations can only occur if that characteristic appears in the population.
The evolution of species has resulted in enormous variation in form and function. Sometimes, evolution gives rise to groups of organisms that become tremendously different from each other. We call two species that evolve in diverse directions from a common point divergent evolution. We can see such divergent evolution in the forms of the reproductive organs of flowering plants which share the same basic anatomies; however, they can look very different as a result of selection in different physical environments and adaptation to different kinds of pollinators.

The flowering plants shown above evolved from a common ancestor. Notice that the dense blazing star (Liatrus spicata) on the left and the purple coneflower (Echinacea purpurea) on the right vary in appearance, yet both share a similar basic morphology. (credit left: a modification of work by Drew Avery; credit right: modification of work by Cory Zanker)
In other cases, similar phenotypes evolve independently in distantly related species. For example, flight has evolved in both bats and insects, and they both have structures we refer to as wings, which are adaptations to flight. However, bat and insect wings have evolved from very different original structures. We call this phenomenon convergent evolution, where similar traits evolve independently in species that do not share a common ancestry. The two species came to the same function, flying, but did so separately from each other.
These physical changes occur over enormous time spans and help explain how evolution occurs. Natural selection acts on individual organisms, which can then shape an entire species. Although natural selection may work in a single generation on an individual, it takes many generations, and therefore often thousands or even millions of years, for an entire species to evolve. It is over these large time spans that life on earth has changed and continues to change.
The Modern Synthesis
The mechanisms of inheritance, genetics, were not understood at the time Darwin and Wallace were developing their idea of natural selection. This lack of understanding was a stumbling block to comprehending many aspects of evolution. In fact, blending inheritance was the predominant (and incorrect) genetic theory of the time, which made it difficult to understand how natural selection might operate. Darwin and Wallace were unaware of the genetics work by Austrian monk Gregor Mendel, which was published in 1866, not long after publication of On the Origin of Species. Mendel’s work was rediscovered in the early twentieth century at which time geneticists were rapidly coming to an understanding of the basics of inheritance. Initially, the newly discovered nature of genes made it difficult for biologists to understand how gradual evolution could occur because each gene was seen as a singular item. But over the next few decades genetics and evolution were integrated in what became known as the modern synthesis—the coherent understanding of the relationship between natural selection and genetics that took shape by the 1940s and is generally accepted today. In sum, the modern synthesis describes how evolutionary pressures, such as natural selection, can affect a population’s genetic makeup, and, in turn, how this can result in the gradual evolution of populations and species. The theory also connects this gradual change of a population over time, called microevolution, with the processes that gave rise to new species and higher taxonomic groups with widely divergent characters, called macroevolution.
Population Genetics
A gene for a particular characteristic may have several variants, or alleles, that code for different traits associated with that characteristic. For example, in the ABO blood type system in humans, three alleles determine the particular blood-type carbohydrate on the surface of red blood cells. Each individual in a population of diploid organisms (organisms, like humans, who have two paired chromosomes, one from each parent) can only carry two alleles for a particular gene, but more than two may be present in the individuals that make up the population. Mendel followed alleles as they were inherited from parent to offspring. In the early twentieth century, biologists began to study what happens to all the alleles in a population in a field of study known as population genetics.
Until now, we have defined evolution as a change in the characteristics of a population of organisms, but behind that phenotypic change is genetic change. In population genetic terms, evolution is defined as a change in the frequency of an allele in a population. Each human has 23 pairs of chromosomes made of strands of DNA. In one of these pairs, each chromosome contains a gene for blood type, the ABO gene. This gene might be any of the three alleles of the ABO gene. Using the ABO system as an example, the frequency of one of the alleles, IA, is the number of copies of that allele divided by all the copies of the ABO gene in the population. For example, a study in Jordan found a frequency of IA to be 26.1 percent.2 The IB, I0 alleles made up 13.4 percent and 60.5 percent of the alleles respectively, and all of the frequencies add up to 100 percent. A change in this frequency over time would constitute evolution in the population.
There are several ways the allele frequencies of a population can change explained below. If a given allele confers a phenotype that allows an individual to have more offspring that survive and reproduce, that allele, by virtue of being inherited by those offspring, will be in greater frequency in the next generation. Since allele frequencies always add up to 100 percent, an increase in the frequency of one allele always means a corresponding decrease in one or more of the other alleles. Highly beneficial alleles may, over a very few generations, become “fixed” in this way, meaning that every individual of the population will carry the allele. Similarly, detrimental alleles may be swiftly eliminated from the gene pool, the sum of all the alleles 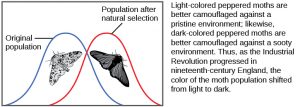 in a population. Part of the study of population genetics is tracking how selective forces change the allele frequencies in a population over time, which can give scientists clues regarding the selective forces that may be operating on a given population. The studies of changes in wing coloration in the peppered moth from mottled white to dark in response to soot-covered tree trunks and then back to mottled white when factories stopped producing so much soot is a classic example of studying evolution in natural populations. As the Industrial Revolution caused trees to darken from soot, darker colored peppered moths were better camouflaged than the lighter colored ones, which caused there to be more of the darker colored moths in the population (see the image above).
in a population. Part of the study of population genetics is tracking how selective forces change the allele frequencies in a population over time, which can give scientists clues regarding the selective forces that may be operating on a given population. The studies of changes in wing coloration in the peppered moth from mottled white to dark in response to soot-covered tree trunks and then back to mottled white when factories stopped producing so much soot is a classic example of studying evolution in natural populations. As the Industrial Revolution caused trees to darken from soot, darker colored peppered moths were better camouflaged than the lighter colored ones, which caused there to be more of the darker colored moths in the population (see the image above).
In the early twentieth century, English mathematician Godfrey Hardy and German physician Wilhelm Weinberg independently provided an explanation for a somewhat counterintuitive concept. Hardy’s original explanation was in response to a misunderstanding as to why a “dominant” allele, one that masks a recessive allele, should not increase in frequency in a population until it eliminated all the other alleles. The question resulted from a common confusion about what “dominant” means, but it forced Hardy, who was not even a biologist, to point out that if there are no factors that affect an allele frequency those frequencies will remain constant from one generation to the next. This principle is now known as the Hardy-Weinberg equilibrium. The theory states that a population’s allele and genotype frequencies are inherently stable—unless some kind of evolutionary force is acting on the population, the population would carry the same alleles in the same proportions generation after generation. Individuals would, as a whole, look essentially the same and this would be unrelated to whether the alleles were dominant or recessive. The four most important evolutionary forces, which will disrupt the equilibrium, are natural selection, mutation, genetic drift, and migration into or out of a population.
Natural Selection
Natural selection has already been discussed. Alleles are expressed in a phenotype. Depending on the environmental conditions, the phenotype confers an advantage or disadvantage to the individual with the phenotype relative to the other phenotypes in the population. If it is an advantage, then that individual will likely have more offspring than individuals with the other phenotypes, and this will mean that the allele behind the phenotype will have greater representation in the next generation. If conditions remain the same, those offspring, which are carrying the same allele, will also benefit. Over time, the allele will increase in frequency in the population.
Mutation
Mutation is a source of new alleles in a population. Mutation is a change in the DNA sequence of the gene. A mutation can change one allele into another, but the net effect is a change in frequency. The change in frequency resulting from mutation is small, so its effect on evolution is small unless it interacts with one of the other factors, such as selection. A mutation may produce an allele that is selected against, selected for, or selectively neutral. Harmful mutations are removed from the population by selection and will generally only be found in very low frequencies equal to the mutation rate. Beneficial mutations will spread through the population through selection, although that initial spread is slow. Whether or not a mutation is beneficial or harmful is determined by whether it helps an organism survive to sexual maturity and reproduce. Mutation is the ultimate source of genetic variation in all populations—new alleles, and, therefore, new genetic variations arise through mutation. Mutation is also random – organisms, species and populations cannot intentionally develop a mutation that is beneficial.
Genetic Drift
Another way a population’s allele frequencies can change is genetic drift, which is simply the effect of chance and is most important in small populations. This process is illustrated in the image at right.  Genetic drift occurs because the alleles in an offspring generation are a random sample of the alleles in the parent generation. Alleles may or may not make it into the next generation due to chance events. For example, consider the simplified example of a population of 10 rabbits in the figure. The dominant allele (A) leads to brown coloration and the recessive allele (a) leads to white coloration. Therefore, any rabbit with an A gene will be brown and only rabbits with two a genes will be white. In the first generation the frequency of the dominant allele (p) equals the frequency of the recessive allele (q). The sum of these two frequencies must add up to one, so each frequency is 0.5. We can find out how many alleles of each type there are by multiplying the frequency (0.5) by the total number of genes (two genes for fur color in each rabbit multiplied by ten rabbits gives 20 genes or 10 of each allele). Imagine a tree falls, killing half of the rabbits at random. In this scenario, the only two white rabbits in the population were killed by chance and therefore unable to reproduce. The remaining five rabbits reproduce to create 10 new rabbits in the second generation, but the frequency of the a allele is now only 0.3. In this generation, another tree falls and kills 8 rabbits, leaving only two rabbits to reproduce. Both happen to have two copies of the dominant allele (A) and so by the third generation, the recessive allele (a) is lost. The two falling trees is obviously improbable, but the key for genetic drift is that the loss of the allele is due to chance. If the white rabbits were easier for predators to see and therefore killed at a greater rate decreasing the frequency of the a allele in future generations, that would be an example of natural selection, not genetic drift.
Genetic drift occurs because the alleles in an offspring generation are a random sample of the alleles in the parent generation. Alleles may or may not make it into the next generation due to chance events. For example, consider the simplified example of a population of 10 rabbits in the figure. The dominant allele (A) leads to brown coloration and the recessive allele (a) leads to white coloration. Therefore, any rabbit with an A gene will be brown and only rabbits with two a genes will be white. In the first generation the frequency of the dominant allele (p) equals the frequency of the recessive allele (q). The sum of these two frequencies must add up to one, so each frequency is 0.5. We can find out how many alleles of each type there are by multiplying the frequency (0.5) by the total number of genes (two genes for fur color in each rabbit multiplied by ten rabbits gives 20 genes or 10 of each allele). Imagine a tree falls, killing half of the rabbits at random. In this scenario, the only two white rabbits in the population were killed by chance and therefore unable to reproduce. The remaining five rabbits reproduce to create 10 new rabbits in the second generation, but the frequency of the a allele is now only 0.3. In this generation, another tree falls and kills 8 rabbits, leaving only two rabbits to reproduce. Both happen to have two copies of the dominant allele (A) and so by the third generation, the recessive allele (a) is lost. The two falling trees is obviously improbable, but the key for genetic drift is that the loss of the allele is due to chance. If the white rabbits were easier for predators to see and therefore killed at a greater rate decreasing the frequency of the a allele in future generations, that would be an example of natural selection, not genetic drift.
In our example above, a natural or human-caused event randomly killed a large portion of the population. This is known as a genetic bottleneck and can result in a large portion of the genome suddenly being wiped out. This concept is illustrated on the right. In one fell swoop,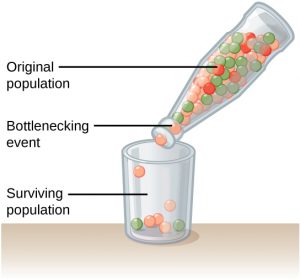 the genetic structure of the survivors becomes the genetic structure of the entire population, which may be very different from the pre-disaster population. In particular, rare alleles are likely to be lost in a genetic bottleneck. The disaster must be one that kills for reasons unrelated to the organism’s traits, such as a hurricane or lava flow. A mass killing caused by unusually cold temperatures at night, is likely to affect individuals differently depending on the alleles they possess that confer cold hardiness.
the genetic structure of the survivors becomes the genetic structure of the entire population, which may be very different from the pre-disaster population. In particular, rare alleles are likely to be lost in a genetic bottleneck. The disaster must be one that kills for reasons unrelated to the organism’s traits, such as a hurricane or lava flow. A mass killing caused by unusually cold temperatures at night, is likely to affect individuals differently depending on the alleles they possess that confer cold hardiness.
Another scenario in which populations might experience a strong influence of genetic drift is if some portion of the population leaves to start a new population in a new location, or if a population gets divided by a physical barrier of some kind. In this situation, those individuals are unlikely to be representative of the entire population which results in the founder effect. The founder effect occurs when the genetic structure matches that of the new population’s founding fathers and mothers. The founder effect is believed to have been a key factor in the genetic history of the Afrikaner population of Dutch settlers in South Africa, as evidenced by mutations that are common in Afrikaners but rare in most other populations. This is likely due to a higher-than-normal proportion of the founding colonists, which were a small sample of the original population, carrying these mutations. As a result, the population expresses unusually high incidences of Huntington’s disease (HD) and Fanconi anemia (FA), a genetic disorder known to cause bone marrow and congenital abnormalities, and even cancer.3
CONCEPTS IN ACTION
Visit this site to learn more about genetic drift and to run simulations of allele changes caused by drift.
Gene Flow and Migration
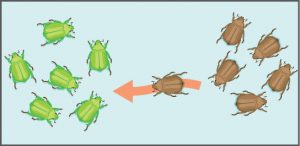 Another important evolutionary force is gene flow, or the flow of alleles in and out of a population resulting from the migration of individuals or seeds. In the example shown to the left, the brown allele is introduced into the green population. While some populations are fairly stable, others experience more flux. Many plants, for example, send their seeds far and wide, by wind or in the guts of animals; these seeds may introduce alleles common in the source population to a new population in which they are rare.
Another important evolutionary force is gene flow, or the flow of alleles in and out of a population resulting from the migration of individuals or seeds. In the example shown to the left, the brown allele is introduced into the green population. While some populations are fairly stable, others experience more flux. Many plants, for example, send their seeds far and wide, by wind or in the guts of animals; these seeds may introduce alleles common in the source population to a new population in which they are rare.
Species and the Ability to Reproduce
Although all life on earth shares various genetic similarities, only certain organisms combine genetic information by sexual reproduction and have offspring that can then successfully reproduce. Scientists call such organisms members of the same biological species.
A species is a group of individual organisms that interbreed and produce fertile, viable offspring. According to this definition, one species is distinguished from another when, in nature, it is not possible for matings between individuals from each species to produce fertile offspring. In reality, the definition of a species is not so clean-cut. For example, some species, like the carrion crow and the hooded crow occasionally interbreed and produce viable offspring, called hybrids. While most hybrids are sterile, the combined offspring of a carrion crow and a hooded crow is often fertile. Scientists now consider these birds evolving species, descendants of a common ancestor that are becoming distinct species. Group of organisms in close proximity in space or time can also challenge this definition of species by partial interbreeding. For example, many, but not all, of the subspecies of Ensatina salamanders can interbreed, and all of the subspecies can interbreed with some of the other subspecies. Like the crows, scientists consider this evidence of evolution in action. Finally, for organisms that reproduce asexually, the biological definition of species is inapplicable and species are generally defined by genetic similarity.
Populations of species share a gene pool: a collection of all the gene variants in the species. Again, the basis to any changes in a group or population of organisms must be genetic for this is the only way to share and pass on traits. When variations occur within a species, they can only pass to the next generation along two main pathways: asexual reproduction or sexual reproduction. The change will pass on asexually simply if the reproducing cell possesses the changed trait. For the changed trait to pass on by sexual reproduction, a gamete, such as a sperm or egg cell, must possess the changed trait. In other words, sexually-reproducing organisms can experience several genetic changes in their body cells, but if these changes do not occur in a sperm or egg cell, the changed trait will never reach the next generation. Only heritable traits can evolve. Therefore, reproduction plays a paramount role for genetic change to take root in a population or species. In short, organisms must be able to reproduce with each other to pass new traits to offspring.
Given the extraordinary diversity of life on the planet there must be mechanisms for speciation: the formation of two species from one original species. Darwin envisioned this process as a branching event and diagrammed the process in the only illustration  in On the Origin of Species (the left panel of the image at right). Compare this illustration to the diagram of elephant evolution (the right panel of the image at right), which shows that as one species changes over time, it branches to form more than one new species, repeatedly, as long as the population survives or until the organism becomes extinct. Darwin’s diagram shows many similarities to phylogenetic charts that today illustrate the relationships of species.
in On the Origin of Species (the left panel of the image at right). Compare this illustration to the diagram of elephant evolution (the right panel of the image at right), which shows that as one species changes over time, it branches to form more than one new species, repeatedly, as long as the population survives or until the organism becomes extinct. Darwin’s diagram shows many similarities to phylogenetic charts that today illustrate the relationships of species.
For speciation to occur, two new populations must form from one original population and they must evolve in such a way that it becomes impossible for individuals from the two new populations to interbreed. Biologists have proposed mechanisms by which this could occur that fall into two broad categories. Allopatric speciation (allo- = “other”; -patric = “homeland”) involves geographic separation of populations from a parent species and subsequent evolution. Sympatric speciation (sym- = “same”; -patric = “homeland”) involves speciation occurring within a parent species remaining in one location.
Biologists think of speciation events as the splitting of one ancestral species into two descendant species. There is no reason why more than two species might not form at one time except that it is less likely and we can conceptualize multiple events as single splits occurring close in time.
Allopatric Speciation
A geographically continuous population has a gene pool that is relatively homogeneous. Gene flow, the movement of alleles across a species’ range, is relatively free because individuals can move and then mate with individuals in their new location. Thus, an allele’s frequency at one end of a distribution will be similar to the allele’s frequency at the other end. When populations become geographically discontinuous, it prevents alleles’ free-flow. When that separation lasts for a period of time, the two populations are able to evolve along different trajectories. Thus, their allele frequencies at numerous genetic loci gradually become increasingly different as new alleles independently arise by mutation in each population. Typically, environmental conditions, such as climate, resources, predators, and competitors for the two populations will differ causing natural selection to favor divergent adaptations in each group.
Isolation of populations leading to allopatric speciation can occur in a variety of ways: a river forming a new branch, erosion creating a new valley, a group of organisms traveling to a new location without the ability to return, or seeds floating over the ocean to an island. The nature of the geographic separation necessary to isolate populations depends entirely on the organism’s biology and its potential for dispersal. If two flying insect populations took up residence in separate nearby valleys, chances are, individuals from each population would fly back and forth continuing gene flow. However, if a new lake divided two rodent populations continued gene flow would be unlikely; therefore, speciation would be more likely.
Additionally, scientists have found that the further the distance between two groups that once were the same species, the more likely it is that speciation will occur. This seems logical because as the distance increases, the various environmental factors would likely have less in common than locations in close proximity. This variation will not lead to new species unless some barrier to successful reproduction also develops.
Adaptive Radiation
In some cases, a population of one species disperses throughout an area, and each finds a distinct niche or isolated habitat. Over time, the varied demands of their new lifestyles lead to multiple speciation events originating from a single species. We call this adaptive radiation because many adaptations evolve from a single point of origin; thus, causing the species to radiate into several new ones. Island archipelagos like the Hawaiian Islands provide an ideal context for adaptive radiation events because water surrounds each island which leads to geographical isolation for many organisms. The Hawaiian honeycreeper illustrates one example of adaptive radiation. From a single species, the founder species, numerous species have evolved, including the six shown below.
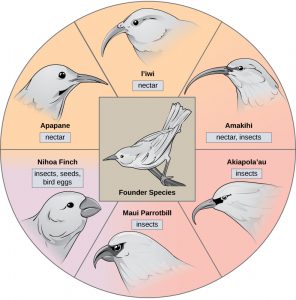
Notice the differences in the species’ beaks above. Evolution in response to natural selection based on specific food sources in each new habitat led to evolution of a different beak suited to the specific food source. The seed-eating bird has a thicker, stronger beak which is suited to break hard nuts. The nectar-eating birds have long beaks to dip into flowers to reach the nectar. The insect-eating birds have beaks like swords, appropriate for stabbing and impaling insects. Darwin’s finches are another example of adaptive radiation in an archipelago.
LINK TO LEARNING
Watch this video to see how scientists use evidence to understand how birds evolved.
Sympatric Speciation
Can divergence occur if no physical barriers are in place to separate individuals who continue to live and reproduce in the same habitat? The answer is yes. We call the process of speciation within the same space sympatric. The prefix “sym” means same, so “sympatric” means “same homeland” in contrast to “allopatric” meaning “other homeland.” Scientists have proposed and studied many mechanisms.
One form of sympatric speciation can begin with a serious chromosomal error during cell division. In a normal cell division event, chromosomes replicate, pair up, and then separate so that each new cell has the same number of chromosomes. However, sometimes 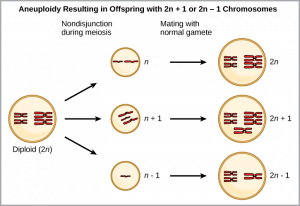 the pairs separate and the end cell product has too many or too few individual chromosomes in a condition that we call aneuploidy. Aneuploidy results when the gametes have too many or too few chromosomes due to nondisjunction during meiosis. In the example at right, the resulting offspring will have 2n+1 or 2n-1 chromosomes. This individual often cannot sexually reproduce with other members of the same species, but may be able to self-pollinate, reproduce asexually or reproduce with other individuals from the same species with aneuploidy. In addition, an organisms with aneuploidy can sometimes reproduce viable offspring with an organism of a different species. This type of process created the cultivated form of wheat. This type of speciation is far more common in plant species than in animal species.
the pairs separate and the end cell product has too many or too few individual chromosomes in a condition that we call aneuploidy. Aneuploidy results when the gametes have too many or too few chromosomes due to nondisjunction during meiosis. In the example at right, the resulting offspring will have 2n+1 or 2n-1 chromosomes. This individual often cannot sexually reproduce with other members of the same species, but may be able to self-pollinate, reproduce asexually or reproduce with other individuals from the same species with aneuploidy. In addition, an organisms with aneuploidy can sometimes reproduce viable offspring with an organism of a different species. This type of process created the cultivated form of wheat. This type of speciation is far more common in plant species than in animal species.
Habitat Influence on Speciation
Sympatric speciation may also take place in ways other than polyploidy. For example, consider a fish species that lives in a lake. As the population grows, competition for food increases. Under pressure to find food, suppose that a group of these fish had the genetic flexibility to discover and feed off another resource that other fish did not use. What if this new food source was located at a different depth of the lake? Over time, those feeding on the second food source would interact more with each other than the other fish; therefore, they would breed together as well. Offspring of these fish would likely behave as their parents: feeding and living in the same area and keeping separate from the original population. If this group of fish continued to remain separate from the first population, eventually sympatric speciation might occur as more genetic differences accumulated between them.
 This scenario does play out in nature, as do others that lead to reproductive isolation. One such place is Lake Victoria in Africa, famous for its sympatric speciation of cichlid fish. Researchers have found hundreds of sympatric speciation events in these fish, which have not only happened in great number, but also over a short period of time. The figure on the left shows this type of speciation among a cichlid fish population in Nicaragua. Lake Apoyeque, a crater lake, is 1800 years old, but genetic evidence indicates that a single population of cichlid fish populated the lake only 100 years ago. Nevertheless, two populations with distinct morphologies and diets now exist in the lake, and scientists believe these populations may be in an early stage of speciation.
This scenario does play out in nature, as do others that lead to reproductive isolation. One such place is Lake Victoria in Africa, famous for its sympatric speciation of cichlid fish. Researchers have found hundreds of sympatric speciation events in these fish, which have not only happened in great number, but also over a short period of time. The figure on the left shows this type of speciation among a cichlid fish population in Nicaragua. Lake Apoyeque, a crater lake, is 1800 years old, but genetic evidence indicates that a single population of cichlid fish populated the lake only 100 years ago. Nevertheless, two populations with distinct morphologies and diets now exist in the lake, and scientists believe these populations may be in an early stage of speciation.
Reproductive Isolation
Given enough time, the genetic and phenotypic divergence between populations will affect characters that influence reproduction: if individuals of the two populations were brought together, mating would be less likely, but if mating occurred, offspring would be nonviable or infertile. For organisms that reproduce sexually, this reproductive isolation is necessary to maintain distinct species. Many types of diverging characteristics may affect the reproductive isolation, the ability to interbreed, of the two populations.
Reproductive isolation can take place in a variety of ways. Scientists organize them into two groups: prezygotic barriers and postzygotic barriers. Recall that a zygote is a fertilized egg: the first cell of an organism’s development that reproduces sexually. Therefore, a prezygotic barrier is a mechanism that blocks reproduction from taking place. This includes barriers that prevent fertilization when organisms attempt reproduction. A postzygotic barrier occurs after zygote formation. This includes organisms that don’t survive the embryonic stage and those that are born sterile.
Some types of prezygotic barriers prevent reproduction entirely. Many organisms only reproduce at certain  times of the year, often just annually. Differences in breeding schedules, which we call temporal isolation, can act as a form of reproductive isolation. For example, two frog species at right inhabit the same area, but one (Rana aurora) reproduces from January to March; whereas, the other (Rana boylii) reproduces from March to May (Photo credits left: modification of work by Mark R. Jennings, USFWS; right: modification of work by Alessandro Catenazzi).
times of the year, often just annually. Differences in breeding schedules, which we call temporal isolation, can act as a form of reproductive isolation. For example, two frog species at right inhabit the same area, but one (Rana aurora) reproduces from January to March; whereas, the other (Rana boylii) reproduces from March to May (Photo credits left: modification of work by Mark R. Jennings, USFWS; right: modification of work by Alessandro Catenazzi).
Behavioral isolation occurs when the presence or absence of a specific behavior prevents reproduction. For example, male fireflies use specific light patterns to attract females. Various firefly species display their lights differently. If a male of one species tried to attract the female of another, she would not recognize the light pattern and would not mate with the male.
Other prezygotic barriers work when differences in their gamete cells (eggs and sperm) prevent fertilization from taking place. We call this a gametic barrier. Similarly, in some cases closely related organisms try to mate, but their reproductive structures simply do not fit together. For example, damselfly males of different species have differently shaped reproductive organs as shown in the image below. If one species tries to mate with the female of another, their body parts simply do not fit together. Reproductive organ incompatibility keeps the species reproductively isolated. 
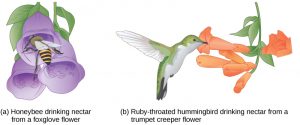
In plants, certain structures aimed to attract one type of pollinator simultaneously prevent a different pollinator from accessing the pollen. The tunnel through which an animal must access nectar can vary widely in length and diameter, which prevents the plant from cross-pollinating with a different species. For example, the wide foxglove flower (left) is adapted for pollination by bees, while the long, tube-shaped trumpet creeper flower (right) is adapted for pollination by hummingbirds.
When fertilization takes place and a zygote forms, postzygotic barriers can prevent reproduction. Hybrid individuals in many cases cannot form normally and simply do not survive past the embryonic stages. We call this hybrid inviability because the hybrid organisms simply are not viable. In another postzygotic situation, reproduction leads to hybrid birth and growth that is sterile. Therefore, the organisms are unable to reproduce offspring of their own. We call this hybrid sterility.
Evidence of Evolution
The evidence for evolution is compelling and extensive. Looking at every level of organization in living systems, biologists see the signature of past and present evolution. Darwin dedicated a large portion of his book, On the Origin of Species, to identifying patterns in nature that were consistent with evolution, and since Darwin, our understanding has become clearer and broader.
Fossils
 Fossils provide solid evidence that organisms from the past are not the same as those today, and fossils show a progression of evolution. Scientists determine the age of fossils and categorize them from all over the world to determine when the organisms lived relative to each other. The resulting fossil record tells the story of the past and shows the evolution of form over millions of years. For example, scientists have recovered highly detailed records showing the evolution of humans and horses. The image to the left shows a display with fossil hominids are arranged from oldest (bottom) to newest (top) on the left. As hominids evolved, the skull’s shape changed. The right half of the image shows an artist’s rendition of extinct species of the genus Equus which reveals that these ancient species resembled the modern horse (Equus ferus) but varied in size.
Fossils provide solid evidence that organisms from the past are not the same as those today, and fossils show a progression of evolution. Scientists determine the age of fossils and categorize them from all over the world to determine when the organisms lived relative to each other. The resulting fossil record tells the story of the past and shows the evolution of form over millions of years. For example, scientists have recovered highly detailed records showing the evolution of humans and horses. The image to the left shows a display with fossil hominids are arranged from oldest (bottom) to newest (top) on the left. As hominids evolved, the skull’s shape changed. The right half of the image shows an artist’s rendition of extinct species of the genus Equus which reveals that these ancient species resembled the modern horse (Equus ferus) but varied in size.
Anatomy and Embryology
Another type of evidence for evolution is the presence of structures in organisms that share the same basic form. 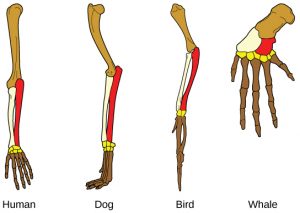 For example, as shown to the right, the bones in human, dog, bird, and whale appendages all share the same overall construction resulting from their origin in a common ancestor’s appendages. Over time, evolution led to changes in the bones’ shapes and sizes different species, but they have maintained the same overall layout. Scientists call these synonymous parts homologous structures.
For example, as shown to the right, the bones in human, dog, bird, and whale appendages all share the same overall construction resulting from their origin in a common ancestor’s appendages. Over time, evolution led to changes in the bones’ shapes and sizes different species, but they have maintained the same overall layout. Scientists call these synonymous parts homologous structures.
Some structures exist in organisms that have no apparent function at all and appear to be residual parts from a past common ancestor. We call these unused structures without function vestigial structures. Some examples of vestigial structures are wings on flightless birds, leaves on some cacti, and hind leg bones in whales. Not all similarities represent homologous structures. Sometimes, similar characteristics occur because of environmental constraints and not due to a close evolutionary relationship. This is called a homoplasy. For example, insects use wings to fly like bats and birds, but the wing structure and embryonic origin are completely different.
This convergence of form in organisms that share similar environments is also evidence of evolution. For example, species of unrelated animals, such as the arctic fox and ptarmigan, living in the arctic region have been selected for seasonal white phenotypes during winter to blend with the snow and ice. These similarities occur not because of common ancestry, but because of similar selection pressures—the benefits of predators not seeing them.
LINK TO LEARNING
Watch this video exploring the bones in the human body.
Biogeography
The geographic distribution of organisms on the planet follows patterns that we can explain best by evolution in conjunction with tectonic plate movement over geological time. Broad groups that evolved before the supercontinent Pangaea broke up (about 200 million years ago) are distributed worldwide. Groups that evolved since the breakup appear uniquely in regions of the planet, such as the unique flora and fauna of northern continents that formed from the supercontinent Laurasia and of the southern continents that formed from the supercontinent Gondwana. The presence of members of the plant family Proteaceae in Australia, southern Africa, and South America was most predominant prior to the southern supercontinent Gondwana breaking up.
Marsupial diversification in Australia and the absence of other mammals reflect Australia’s long isolation. Australia has an abundance of endemic species—species found nowhere else—which is typical of islands whose isolation by expanses of water prevents species migration. Over time, these species diverge evolutionarily into new species that look very different from their ancestors that may exist on the mainland. Australia’s marsupials, the Galápagos’ finches, and many species on the Hawaiian Islands are all unique to their one point of origin, yet they display distant relationships to ancestral species on mainlands. The concept of biogeography is explored further in Chapter 4.
Molecular Biology
Like anatomical structures, the molecular structures of life reflect descent with modification. DNA’s universality reflects evidence of a common ancestor for all of life. In general, the relatedness of groups of organisms is reflected in the similarity of their DNA sequences—exactly the pattern that we would expect from descent and diversification from a common ancestor. DNA sequences have also shed light on some of the mechanisms of evolution. For example, it is clear that the evolution of new functions for proteins commonly occurs after gene duplication events that allow freely modifying one copy by mutation, selection, or drift (changes in a population’s gene pool resulting from chance), while the second copy continues to produce a functional protein.
Misconceptions of Evolution
Although the theory of evolution generated some controversy when Darwin first proposed it, biologists almost universally accepted it, particularly younger biologists, within 20 years after publication of On the Origin of Species. Nevertheless, the theory of evolution is a difficult concept and misconceptions about how it works abound.
LINK TO LEARNING
This site addresses some of the main misconceptions associated with the theory of evolution.
Evolution Is Just a Theory
Critics of the theory of evolution dismiss its importance by purposefully confounding the everyday usage of the word “theory” with the way scientists use the word. As discussed in Chapter 1, a scientific theory is always based on evidence and an accepted scientific theory has has been thoroughly tested and provides an explanation for a set of observations of the natural world. Scientists have a theory of the atom, a theory of gravity, and the theory of relativity, each which describes understood facts about the world. In the same way, the theory of evolution describes facts about the living world. A fundamental theory in science like the theory of evolution has survived significant efforts to discredit it by scientists. In contrast, a “theory” in common vernacular is a word meaning a guess or suggested explanation. When critics of evolution say it is “just a theory,” they are implying that there is little evidence supporting it and that it is still in the process of rigorous testing. This is a mischaracterization.
Individuals Evolve
Evolution is the change in a population’s genetic composition over time, specifically over generations, resulting from differential reproduction of individuals with certain alleles. Individuals do change over their lifetime, obviously, but this is development and involves changes programmed by the set of genes the individual acquired at birth in coordination with the individual’s environment. When thinking about the evolution of a characteristic, it is probably best to think about the change of the average value of the characteristic in the population over time. For example, when natural selection leads to bill-size change in medium ground finches in the Galápagos, this does not mean that individual bills on the finches are changing. If one measures the average bill size among all individuals in the population at one time and then measures them in the population several years later, this average value will be different as a result of evolution. Although some individuals may survive from the first time to the second, they will still have the same bill size; however, there will be many new individuals who contribute to the shift in average bill size.
Evolution Explains the Origin of Life
It is a common misunderstanding that evolution includes an explanation of life’s origins. Conversely, some of the theory’s critics believe that it cannot possibly explain the origin of life. The theory does not try to explain the origin of life. The theory of evolution explains how populations change over time and how life diversifies the origin of species. It does not shed light on the beginnings of life including the origins of the first cells, which define life.
Organisms Evolve on Purpose
Statements such as “organisms evolve in response to a change in an environment” are quite common, but such statements can lead to two types of misunderstandings. First, do not interpret the statement to mean that individual organisms evolve. The statement is shorthand for “a population evolves in response to a changing environment.” However, a second misunderstanding may arise by interpreting the statement to mean that the evolution is somehow intentional. A changed environment results in some individuals in the population, those with particular phenotypes, benefiting and therefore producing proportionately more offspring than other phenotypes. This results in change in the population if the characteristics are genetically determined.
It is also important to understand that the variation that natural selection works on is already in a population and does not arise in response to an environmental change. For example, applying antibiotics to a population of bacteria will, over time, select a population of bacteria that are resistant to antibiotics. The resistance, which a gene causes, did not arise by mutation because of applying the antibiotic. The gene for resistance was already present in the bacteria’s gene pool, likely at a low frequency, or was produced by a random mutation. The antibiotic, which kills the bacterial cells without the resistance gene, strongly selects individuals that are resistant, since these would be the only ones that survived and divided. Experiments have demonstrated that mutations for antibiotic resistance do not arise as a result of antibiotic.
In a larger sense, evolution is not goal directed. Species do not become “better” over time. They simply track their changing environment with adaptations that maximize their reproduction in a particular environment at a particular time. Evolution has no goal of making faster, bigger, more complex, or even smarter species, despite the commonness of this kind of language in popular discourse. What characteristics evolve in a species are a function of the variation present and the environment, both of which are constantly changing in a nondirectional way. A trait that fits in one environment at one time may well be fatal at some point in the future. This holds equally well for insect and human species.
You can find more information on evolution at https://evolution.berkeley.edu/evolibrary/article/evo_01.
- Footnotes
- 1Theodosius Dobzhansky. “Biology, Molecular and Organismic.” American Zoologist4, no. 4 (1964): 449.
- 2Sahar S. Hanania, Dhia S. Hassawi, and Nidal M. Irshaid, “Allele Frequency and Molecular Genotypes of ABO Blood Group System in a Jordanian Population,” Journal of Medical Sciences7 (2007): 51-58, doi:10.3923/jms.2007.51.58
- 3A. J. Tipping et al., “Molecular and Genealogical Evidence for a Founder Effect in Fanconi Anemia Families of the Afrikaner Population of South Africa,” PNAS98, no. 10 (2001): 5734-5739, doi: 10.1073/pnas.091402398.
- Content modified from
- o Authors: Samantha Fowler, Rebecca Roush, James Wise
- o Publisher/website: OpenStax
- o Book title: Concepts of Biology
- o Publication date: Apr 25, 2013
- o Location: Houston, Texas
- o Book URL: https://openstax.org/books/concepts-biology/pages/1-introduction
- Access for free at https://openstax.org/books/biology-2e@16.144/pages/1-introduction
And from
- Use the information below to generate a citation. We recommend using a citation tool such as this one.
- Authors: Mary Ann Clark, Matthew Douglas, Jung Choi
- Publisher/website: OpenStax
- Book title: Biology 2e
- Publication date: Mar 28, 2018
- Location: Houston, Texas
- Book URL: https://openstax.org/books/biology-2e@16.144/pages/1-introduction
- Section URL: https://openstax.org/books/biology-2e@16.144/pages/1-introduction
Change in the genetics of a population through time.
a mechanism for evolution by which traits that make some individuals able to produce more offspring pass these traits on to these offspring making these traits more common in future generations
heritable trait or behavior in an organism that aids in its survival and reproduction in its present environment
process by which groups of organisms independently evolve to similar forms
the changes in a population’s genetic structure (i.e., allele frequency)
a broader scale of evolutionary changes seen over paleontological time
a change in the DNA sequence
Changes in the genetic makeup of a population due to chance
the movement of individuals of a population to a new location; in population genetics it refers to the movement of individuals and their alleles from one population to another, potentially changing allele frequencies in both the old and the new population
A change in the genetic makeup of a population because of a natural or human-caused event that causes a significant reduction in the number of individuals
group of populations that interbreed and produce fertile offspring
formation of a new species
speciation that occurs via geographic separation
speciation that occurs in the same geographic space
condition of a cell having an extra chromosome or missing a chromosome for its species
reproductive isolation mechanism that occurs before zygote formation
reproductive isolation mechanism that occurs after zygote formation
differences in breeding schedules that can act as a form of prezygotic barrier leading to reproductive isolation
situation that occurs when a species is reproductively independent from other species; behavior, location, or reproductive barriers may cause this to happen
type of reproductive isolation that occurs when a specific behavior or lack of one prevents reproduction from taking place
prezygotic barrier occurring when closely related individuals of different species mate, but differences in their gamete cells (eggs and sperm) prevent fertilization from taking place
parallel structures in diverse organisms that have a common ancestor
physical structure present in an organism but that has no apparent function and appears to be from a functional structure in a distant ancestor
A shared characteristic that did not arise from a common ancestor
