
27 Chapter 27 The Reproductive System
By Rajeev Chandra
Motivation.
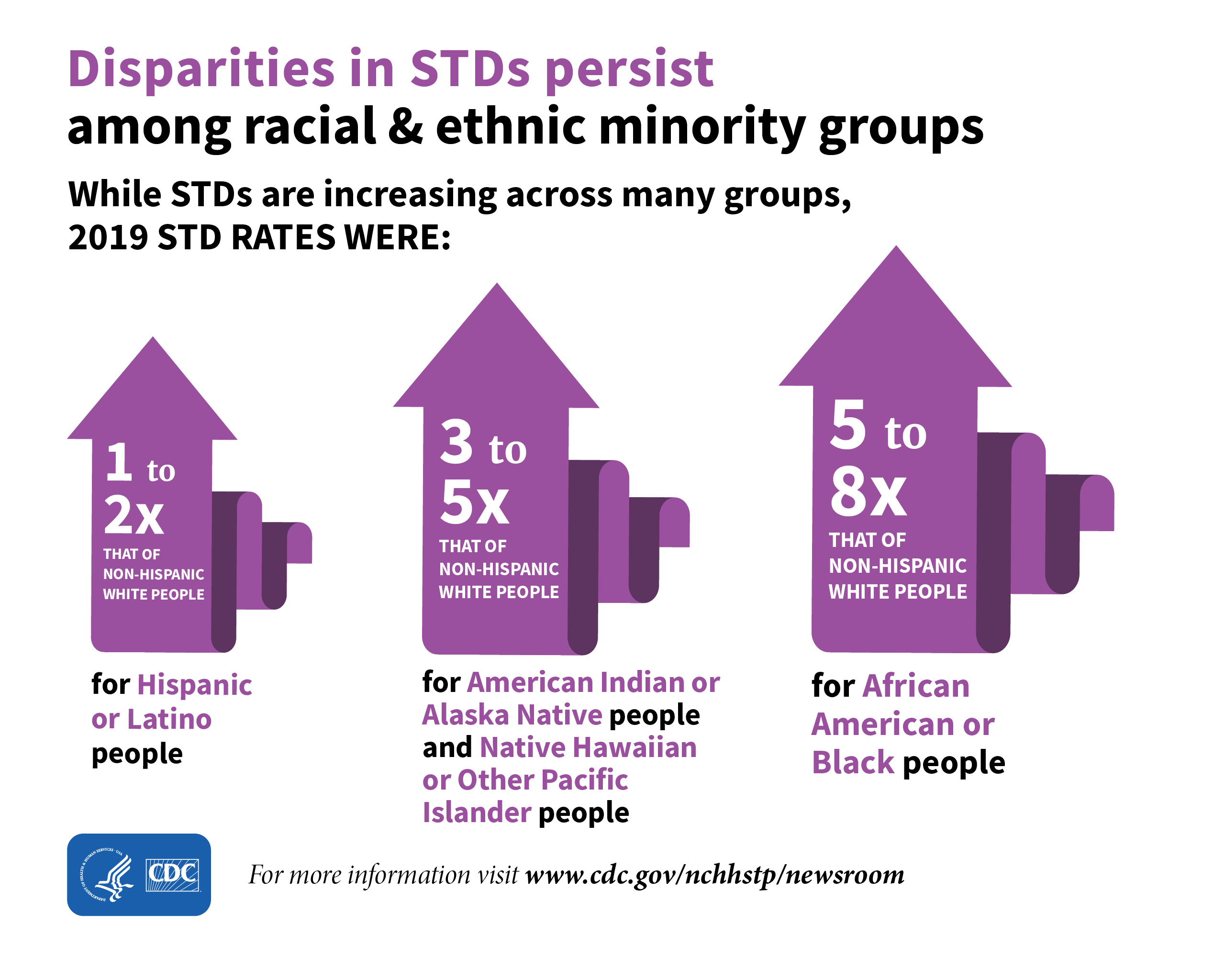
Washington, D.C. has among the highest rates of sexually transmitted diseases (STDs) and unintended pregnancy in the United States. Increasing everyone’s reproductive health knowledge may help address these reproductive health issues. This analysis assessed whether high-risk pregnant African American women in Washington, D.C. who participated in an intervention to reduce behavioral and psychosocial risks had greater reproductive health knowledge than women receiving usual care.
Learning Objectives
Upon completion of the work in this chapter students should be able to:
- Describe female reproductive organ histology and anatomy
- Describe the gross and microscopic anatomy of the male reproductive organs
- Relate the structure of sperm to its function
Background.
Overview of the Female Reproductive System
The female reproductive system functions to produce gametes and reproductive hormones, just like the male reproductive system; however, it also has the additional task of supporting a developing fetus and delivering it to the outside world. Unlike its male counterpart, the female reproductive system is located primarily inside the pelvic cavity (Figure 27.2). Recall that the ovaries are the female gonads and the gamete that is produced is called an oocyte.
Ovaries
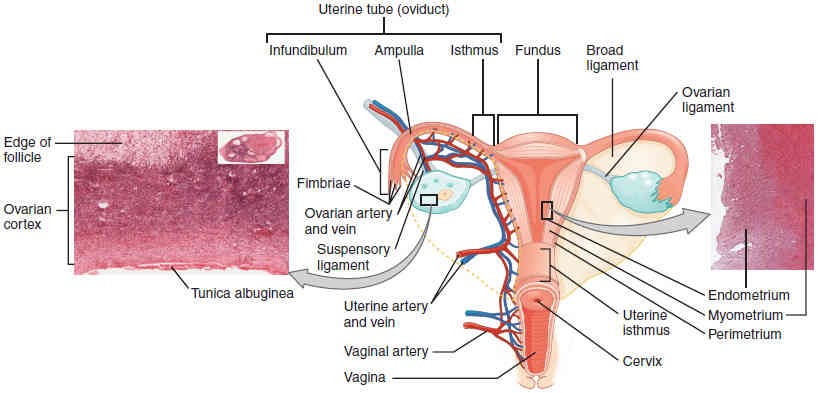
The ovaries are the female gonads (Figure 27.2 and Figure 27.3). Paired ovals, they are each about 2 to 3 cm in length, about the size of an almond. The ovaries are located within the pelvic cavity, and are supported by the mesovarium, an extension of the peritoneum that connects the ovaries to the broad ligament. Extending from the mesovarium itself is the suspensory ligament that contains the ovarian blood and lymphatic vessels. Finally, the ovary itself is attached to the uterus via the ovarian ligament.
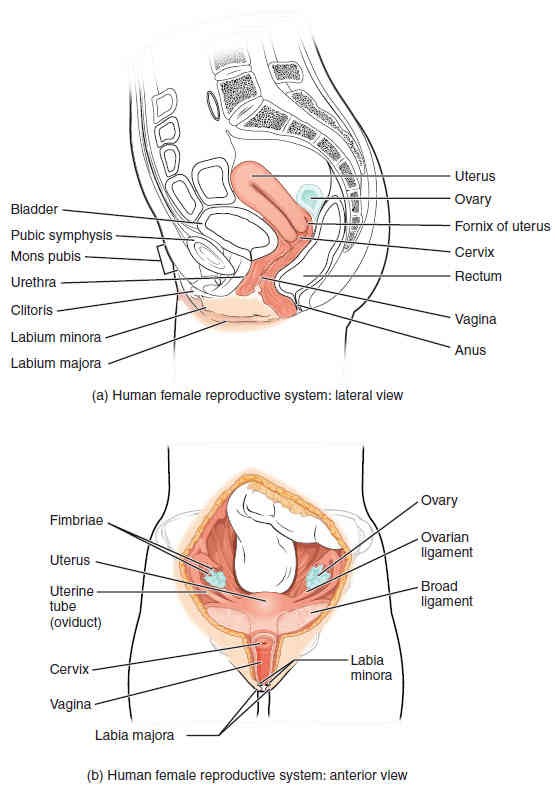
The ovary consists of multiple layers of tissue. The outer-most covering of cuboidal epithelium called the ovarian surface epithelium sits just superficial to a dense connective tissue layer, known as the tunica albuginea. Beneath the tunica albuginea is the cortex, or outer portion, of the organ itself. The cortex is composed of a tissue framework called the ovarian stroma that forms the bulk of the adult ovary. Oocytes develop within the outer layer of this stroma, each surrounded by supporting cells. This grouping of an oocyte and its supporting cells is called a follicle (Figure 27.3). The growth and development of ovarian follicles will be described shortly. Beneath the cortex lies the inner ovarian medulla, where the majority of blood vessels, lymphatic vessels, and the nerves of the ovary are localized to.
The Ovarian Cycle and Oogenesis
The ovarian cycle is a set of predictable changes in a female’s oocytes and ovarian follicles. During a woman’s reproductive years, it is a roughly 28-day cycle that can be correlated with, but is not the same as, the menstrual cycle. The cycle includes two interrelated processes: oogenesis (the production of female gametes) and folliculogenesis (the growth and development of ovarian follicles).
Oogenesis
Gametogenesis in females is called oogenesis. The process begins with ovarian stem cells, or oogonia (pleural: oogonium) (Figure 27.4). Oogonia are formed during fetal development, and divide via mitosis, much like spermatogonia in the testis. Unlike spermatogonia, however, oogonia form primary oocytes in the fetal ovary prior to birth. These primary oocytes are then arrested in prophase of meiosis I, only to resume it years later, beginning at puberty and continuing until the woman is near menopause (the cessation of a woman’s reproductive functions). The number of primary oocytes present in the ovaries declines from one to two million in an infant, to approximately 400,000 at puberty, to zero by the end of menopause.
The unequal cell division of oogenesis produces one to three polar bodies that later degrade, as well as a single haploid ovum, which is produced only if there is penetration of the secondary oocyte by a sperm cell.
The initiation of ovulation, the release of an oocyte from the ovary, marks the transition from puberty into reproductive maturity for women. From the onset of ovulation and throughout a woman’s reproductive years, ovulation occurs approximately once every 28 days. Just prior to ovulation, a surge of luteinizing hormone triggers the resumption of meiosis in a primary oocyte. This initiates the transition from primary to secondary oocyte. However, as you can see in Figure 27.4, this cell division does not result in two identical cells. Instead, the cytoplasm is divided unequally, and one daughter cell is much larger than the other. This larger cell, the secondary oocyte, eventually leaves the ovary during ovulation. The smaller cell, called the first polar body, may or may not complete meiosis and produce second polar bodies; in either case, it eventually disintegrates. Therefore, even though oogenesis produces up to four cells, only one survives.
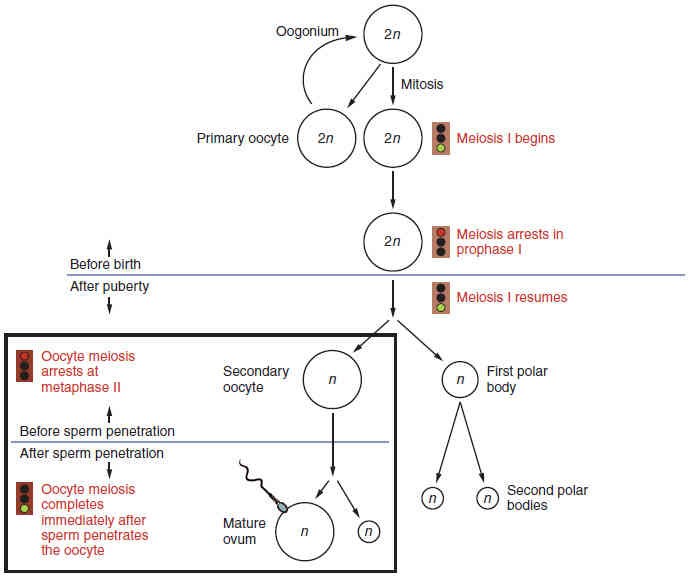
A question still remains though: How does the diploid secondary oocyte become an ovum —the haploid female gamete? Meiosis of a secondary oocyte is completed only if a sperm succeeds in penetrating its barriers. If union of a secondary oocyte and a sperm is successful, only then will meiosis II resume. This fusion will produce one haploid ovum that, at the moment of fertilization by a (haploid) sperm, becomes the first diploid cell of the new offspring (a zygote). Thus, the ovum can be thought of as a brief, transitional, haploid stage between the diploid oocyte and diploid zygote.
The larger amount of cytoplasm contained in the female gamete is used to supply the developing zygote with nutrients during the period between fertilization and implantation into the uterus. Interestingly, sperm contribute only DNA at fertilization —not cytoplasm. Therefore, the cytoplasm and all of the cytoplasmic organelles in the developing embryo are of maternal origin.
Folliculogenesis
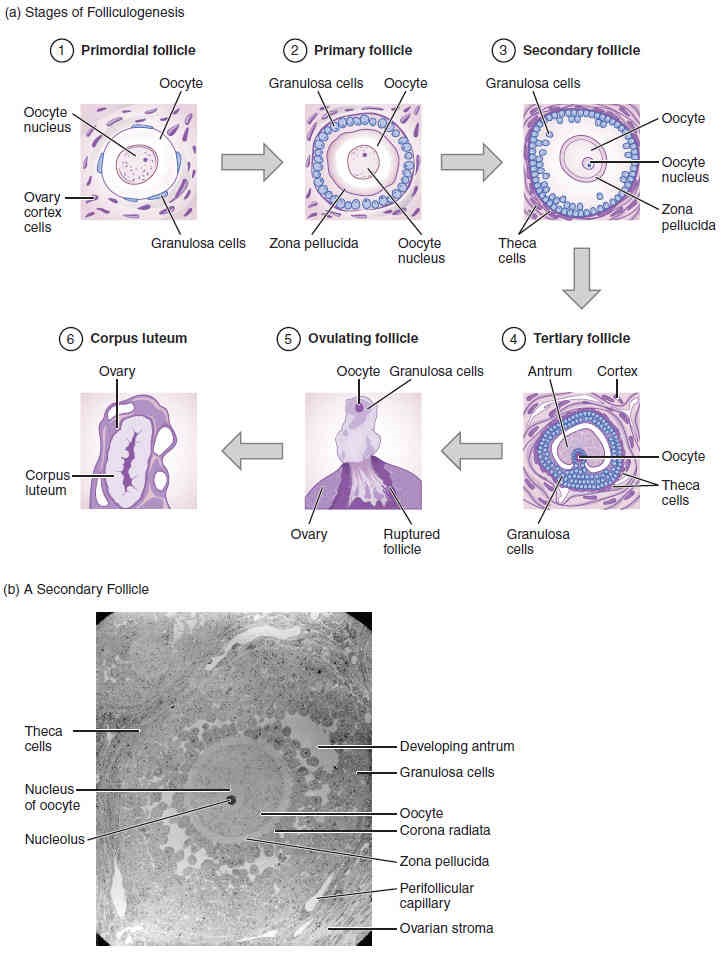
Remember, ovarian follicles are oocytes and their supporting cells. They grow and develop in a process called folliculogenesis, which typically leads to ovulation of one follicle approximately every 28 days, along with death to multiple other follicles. The death of ovarian follicles is called atresia, and can occur at any point during follicular development. Recall that, a female infant at birth will have one to two million oocytes within her ovarian follicles, and that this number declines throughout life until menopause, when no follicles remain. As you’ll see next, follicles progress from primordial, to primary, to secondary and finally tertiary stages prior to ovulation—with the oocyte inside the follicle remaining as a primary oocyte until right before ovulation.
Folliculogenesis begins with follicles in a resting state. These small primordial follicles are present in newborn females and are the prevailing follicle type in the adult ovary (Figure 27.5). Primordial follicles have only a single flat layer of supporting cells, called granulosa cells, that surround the primary oocyte, and they can stay in this resting state for years—some until right before menopause.
After puberty, a few primordial follicles will respond to a recruitment signal each day, and will join a pool of immature growing follicles called primary follicles. Primary follicles start with a single layer of granulosa cells, but the granulosa cells then become active and transition from a flat or squamous shape to a rounded, cuboidal shape as they increase in size and proliferate. As the granulosa cells divide, the follicles—now called secondary follicles (Figure 27.5)—increase in diameter, adding a new outer layer of connective tissue, blood vessels, and theca cells —cells that work with the granulosa cells to produce estrogens. Within the growing secondary follicle, the primary oocyte now secretes a thin acellular membrane called the zona pellucida that will play a critical role in fertilization. A thick fluid, called follicular fluid, that has formed between the granulosa cells also begins to collect into one large pool, the antrum. Follicles in which the antrum has become large and fully formed are considered tertiary follicles (or antral follicles). Several follicles reach the tertiary stage at the same time, and most of these will undergo atresia. The one that does not die will continue to grow and develop until ovulation, when it will expel its secondary oocyte surrounded by several layers of granulosa cells from the ovary. Keep in mind that most follicles don’t make it to this point. In fact, roughly 99 percent of the follicles in the ovary will undergo atresia, which can occur at any stage of folliculogenesis.
Hormonal Control of the Ovarian Cycle
The process of development that we have just described, from primordial follicle to early tertiary follicle, takes approximately two months in humans. The final stages of development of a small cohort of tertiary follicles, ending with ovulation of a secondary oocyte, occur over a course of approximately 28 days. These changes are regulated by many of the same hormones that regulate the male reproductive system, including GnRH, LH, and FSH.
As in men, the hypothalamus produces GnRH, a hormone that signals the anterior pituitary gland to produce the gonadotropins FSH and LH (Figure 27.6). These gonadotropins leave the pituitary and travel through the bloodstream to the ovaries, where they bind to receptors on the granulosa and theca cells of the follicles. FSH stimulates the follicles to grow (hence its name of follicle-stimulating hormone), and the five or six tertiary follicles expand in diameter. The release of LH also stimulates the granulosa and theca cells of the follicles to produce the sex steroid hormone estradiol, a type of estrogen.
This phase of the ovarian cycle, when the tertiary follicles are growing and secreting estrogen, is known as the follicular phase. The more granulosa and theca cells a follicle has (that is, the larger and more developed it is), the more estrogen it will produce in response to LH stimulation. As a result of these large follicles producing large amounts of estrogen, systemic plasma estrogen concentrations increase. Following a classic negative feedback loop, the high concentrations of estrogen will stimulate the hypothalamus and pituitary to reduce the production of GnRH, LH, and FSH. Because the large tertiary follicles require FSH to grow and survive at this point, this decline in FSH caused by negative feedback leads most of them to die (atresia) (Figure 27.6, step 1). Typically, only one follicle, now called the dominant follicle, will survive this reduction in FSH, and this follicle will be the one that releases an oocyte. Scientists have studied many factors that lead to a particular follicle becoming dominant: size, the number of granulosa cells, and the number of FSH receptors on those granulosa cells all contribute to a follicle becoming the one surviving dominant follicle.
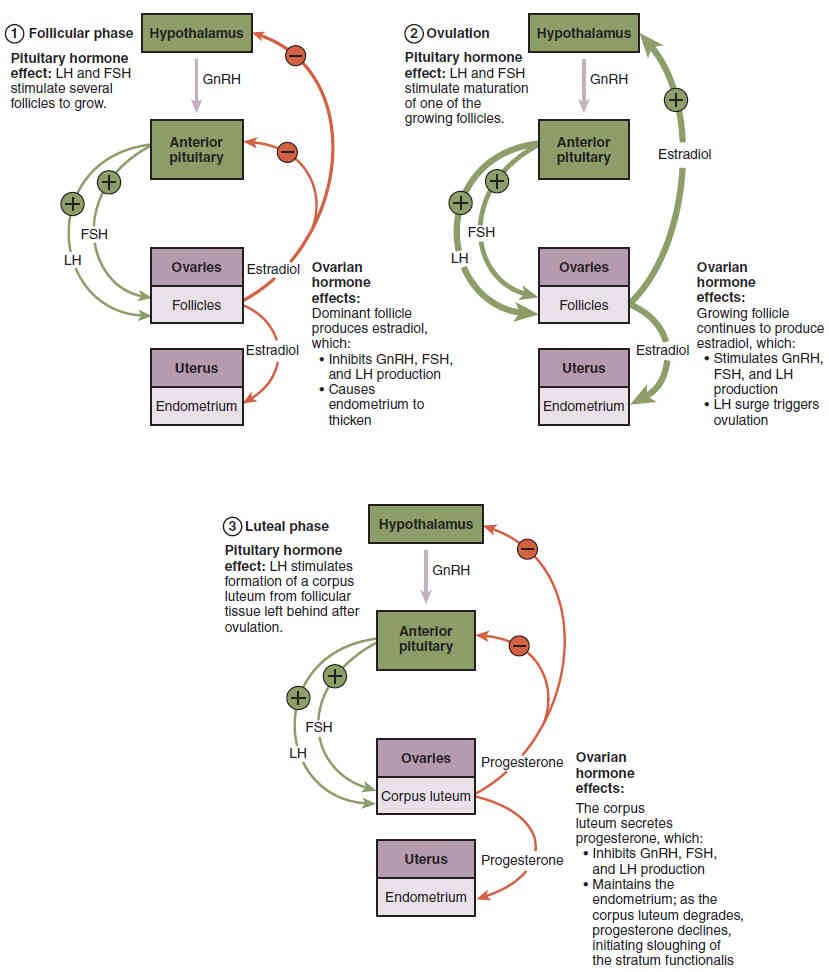
When only the dominant follicle remains in the ovary, it again begins to secrete estrogen. It produces more estrogen than all of the developing follicles did together before the negative feedback occurred. It produces so much estrogen that the normal negative feedback doesn’t occur. Instead, these extremely high concentrations of systemic plasma estrogen trigger a regulatory switch in the anterior pituitary that responds by secreting large amounts of LH and FSH into the bloodstream (Figure 27.6, step 2).
The positive feedback loop by which more estrogen triggers release of more LH and FSH only occurs at this point in the cycle. It is this large burst of LH (called the LH surge) that leads to ovulation of the dominant follicle. The LH surge induces many changes in the dominant follicle, including stimulating the resumption of meiosis of the primary oocyte to a secondary oocyte. As noted earlier, the polar body that results from unequal cell division simply degrades. The LH surge also triggers proteases (enzymes that cleave proteins) to break down structural proteins in the ovary wall on the surface of the bulging dominant follicle. This degradation of the wall, combined with pressure from the large, fluid-filled antrum, results in the expulsion of the oocyte surrounded by granulosa cells into the peritoneal cavity. This release is ovulation.
There is one more important event that occurs in the ovarian cycle. The surge of LH also stimulates a change in the granulosa and theca cells that remain in the follicle after the oocyte has been ovulated. This change is called luteinization and it transforms the collapsed follicle into a new endocrine structure called the corpus luteum, a term meaning “yellowish body” (Figure 27.5). Instead of estrogen, the luteinized granulosa and theca cells of the corpus luteum begin to produce large amounts of the sex steroid hormone progesterone, a hormone that is critical for the establishment and maintenance of pregnancy. Progesterone triggers negative feedback at the hypothalamus and pituitary, which keeps GnRH, LH, and FSH secretions low, so no new dominant follicles develop at this time. This post-ovulatory phase of progesterone secretion is known as the luteal phase of the ovarian cycle (Figure 27.6, step 3). If pregnancy does not occur within 10 to 12 days, the corpus luteum will stop secreting progesterone and degrade into the corpus albicans, a nonfunctional “whitish body” that will degenerate in the ovary over a period of several months. During this time of reduced progesterone secretion, FSH and LH are once again stimulated, and the follicular phase begins again with a new cohort of early tertiary follicles beginning to grow and secrete estrogen.
Uterine (Fallopian) Tubes
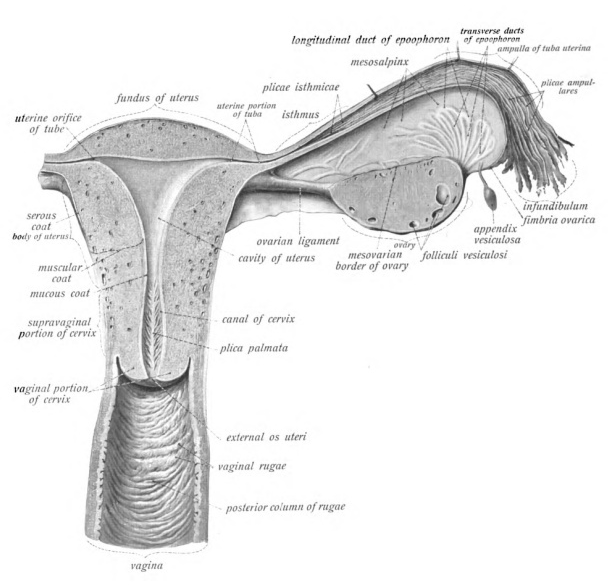
The uterine tubes (also called fallopian tubes or oviducts) serve as the conduit of the oocyte from the ovary to the uterus (Figure 27.3). Each of the two uterine tubes is close to, but not directly connected to, the ovary and each is divided into sections. The isthmus is the narrow medial end of each uterine tube that is connected to the uterus. The wide distal infundibulum flares out with slender, finger-like projections called fimbriae. The middle region of the tube, called the ampulla, is where fertilization often occurs. The uterine tubes also have three layers of tissue: an outer serosa, a middle smooth muscle layer, and an inner mucosal layer. In addition to its mucus-secreting cells, the inner mucosa contains ciliated cells that beat in the direction of the uterus, producing a current that will be critical to move the oocyte.
Following ovulation, the secondary oocyte surrounded by a few granulosa cells is released into the peritoneal cavity. The nearby uterine tube, either left or right, receives the oocyte. Unlike sperm, oocytes lack flagella, and therefore cannot move on their own. So how do they travel into the uterine tube and toward the uterus? High concentrations of estrogen that occur around the time of ovulation induce contractions of the smooth muscle along the length of the uterine tube. These contractions occur every 4 to 8 seconds, and the result is a coordinated movement that sweeps the surface of the ovary and the pelvic cavity. As a result of these mechanisms, the oocyte–granulosa cell complex is pulled into the interior of the tube. Once inside, the muscular contractions and beating cilia move the oocyte slowly toward the uterus. When fertilization does occur, sperm typically meet the egg while it is still moving through the ampulla.
Uterus
The uterus is the muscular organ that nourishes and supports the growing embryo (Figure 27.3). Its average size is approximately 5 cm wide by 7 cm long when a female is not pregnant. It has three sections: the portion of the uterus superior to the opening of the uterine tubes is called the fundus, the middle section of the uterus is called the body or corpus, and the cervix is the narrow inferior portion of the uterus that projects into the vagina.
The wall of the uterus is made up of three layers (Figure 27.3 and Figure 27.7). The most superficial layer is the serous membrane, or perimetrium, which consists of epithelial tissue that covers the exterior portion of the uterus. The middle layer, or myometrium, is a thick layer of smooth muscle responsible for uterine contractions. Most of the uterus is myometrial tissue, and the muscle fibers run horizontally, vertically, and diagonally, allowing the powerful contractions that occur during labor and the less powerful contractions (or cramps) that help to expel menstrual blood during a woman’s period.
The innermost layer of the uterus is called the endometrium. Structurally, the endometrium consists of two layers: the stratum basalis (basal layer) and the stratum functionalis (functional layer). The stratum basalis layer lies adjacent to the myometrium; this layer does not shed during menses. In contrast, the thicker stratum functionalis layer contains the glandular endothelial tissues that line the uterine lumen. It is the stratum functionalis that grows and thickens in response to increased levels of estrogen and progesterone. In the luteal phase of the menstrual cycle, special branches off of the uterine artery called spiral arteries supply the thickened stratum functionalis (Figure 27.7). This inner functional layer provides the proper site of implantation for a fertilized egg, and—should fertilization not occur—it is only the functional layer of the endometrium that sheds during menstruation.
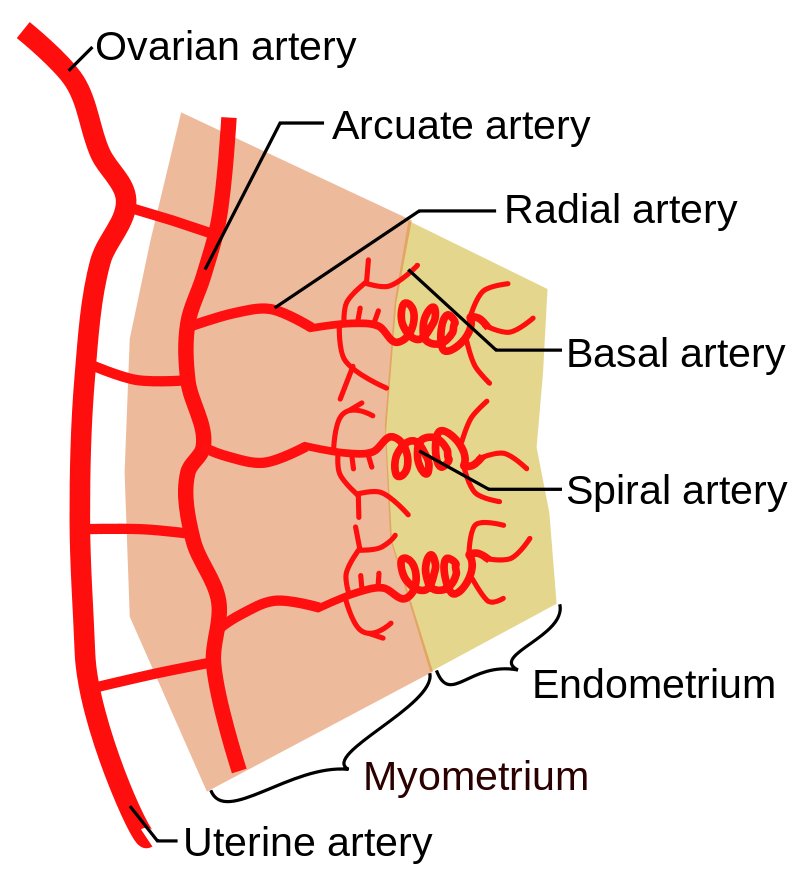
Figure 27.7 Layers and arterial vasculature of the uterus. The wall of the uterus consists of 3 layers: the outer perimetrium (not shown), the myometrium, and the endometrium. Credit: Mikael Häggström, Wikimedia Commons, license Public Domain.
Ligaments
Several ligaments maintain the position of the uterus within the abdominopelvic cavity (Figure 27.3). The broad ligament is a fold of peritoneum that serves as a primary support for the uterus, extending laterally from both sides of the uterus and attaching it to the pelvic wall. The round ligament attaches to the uterus near the uterine tubes, and extends to the labia majora.
Vagina
The vagina, shown at the bottom of Figure 27.2 and in Figure 27.8, is a muscular canal (approximately 10 cm long) that serves as the entrance to the reproductive tract. It also serves as the exit from the uterus during menses and childbirth. The outer walls of the anterior and posterior vagina are formed into longitudinal columns, or ridges, and the superior portion of the vagina—called the fornix—meets the protruding uterine cervix. The walls of the vagina are lined with an outer, fibrous adventitia, a middle layer of smooth muscle, and an inner mucous membrane with transverse folds called rugae. Together, the middle and inner layers allow the expansion of the vagina to accommodate intercourse and childbirth. A thin, perforated hymen can partially surround the opening to the vaginal orifice (opening).
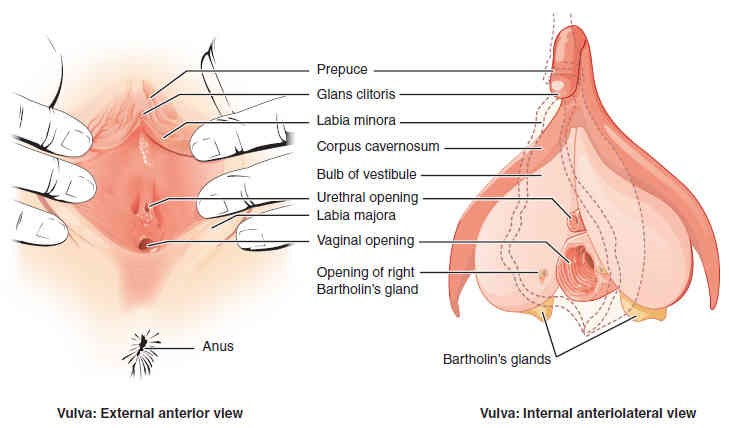
External Genitalia
The external female reproductive structures are referred to collectively as the vulva (Figures 27.2 and 27.9) and they include the structures that will be discussed next. The mons pubis is a pad of fat that is located anteriorly, over the pubic bone. After puberty, it becomes covered in pubic hair. The labia majora (labia = “lips”; majora = “larger”) are folds of hair-covered skin that begin just posterior to the mons pubis. The thinner and more pigmented labia minora (labia = “lips”; minora = “smaller”) extend medial to the labia majora and the space between labia minora is known as the vestibule. Although they naturally vary in shape and size from woman to woman, the labia minora serve to protect the female urethra and the entrance to the female reproductive tract.
The superior, anterior portions of the labia minora come together to encircle the clitoris (or glans clitoris), an organ that originates from the same cells as the glans penis, and has abundant nerves that make it important in sexual sensation and orgasm. The hymen is a thin membrane that sometimes partially covers the entrance to the vagina. The vaginal opening, also known as the vaginal orifice, is located between the opening of the urethra and the anus. It is flanked by outlets to the Bartholin’s glands (or greater vestibular glands) .
The Breast
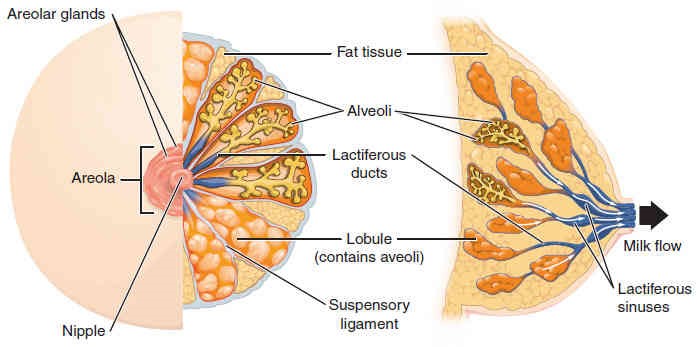
Whereas the breasts are located far from the other female reproductive organs, they are considered accessory organs of the female reproductive system. The function of the breasts is to supply milk to an infant in a process called lactation. The external features of the breast include a nipple surrounded by a pigmented areola (Figure 27.10), whose coloration may deepen during pregnancy. The areolar region is characterized by small, raised areolar glands that secrete lubricating fluid during lactation to protect the nipple from chafing.
Internally, breast milk is produced by the mammary glands, which are modified sweat glands. The milk itself exits the breast through the nipple via 15 to 20 lactiferous ducts that open on the surface of the nipple. These lactiferous ducts each extend to a lactiferous sinus that connects to a glandular lobe (lobule) within the breast itself that contains groups of milksecreting cells in clusters called alveoli (Figure 27.10). Once milk is made in the alveoli, stimulated myoepithelial cells that surround the alveoli contract to push the milk to the lactiferous sinuses. From here, a baby can draw milk through the lactiferous ducts by suckling. The lobules themselves are surrounded by fat tissue, which determines the size of the breast; breast size differs between individuals and does not affect the amount of milk produced. Supporting the breasts are multiple bands of connective tissue called suspensory ligaments that connect the breast tissue to the dermis of the overlying skin.
During the normal hormonal fluctuations in the menstrual cycle, breast tissue responds to changing levels of estrogen and progesterone, which can lead to swelling and breast tenderness in some individuals, especially during the secretory phase. If pregnancy occurs, the increase in hormones leads to further development of the mammary tissue and enlargement of the breasts.
Overview of the Male Reproductive System
The function of the male reproductive system is to produce male gametes, known as sperm, to transfer these to the female reproductive tract, and to secrete the hormones that support male reproductive physiology. The paired gonads, or gamete-producing structures, are the testes (singular, testis) and they are a crucial component of the male’s reproductive system. While the testes produce both sperm and androgens, several accessory organs and ducts aid in the process of sperm maturation and transport of the sperm and other seminal components to the penis, which delivers sperm to the female reproductive tract.
The structures of the male reproductive system include the testes, the epididymis, and the penis, as well as the ducts and glands that produce and carry semen (Figure 27.11).
Scrotum and Testes
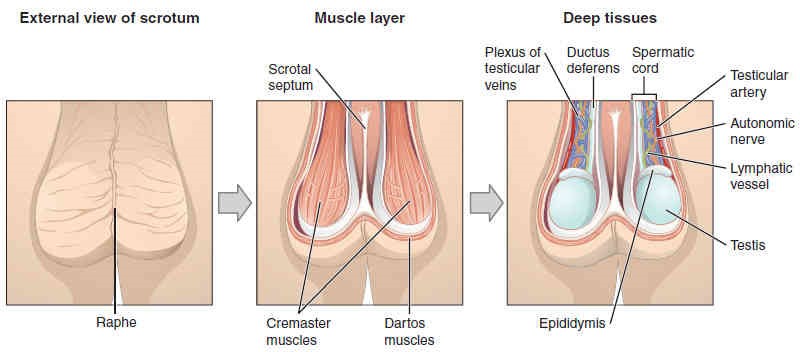
The testes (singular, testis) are located in a skin-covered, highly pigmented, muscular sack called the scrotum. The scrotum extends from the body behind the penis (Figure 27.11). This location is important to sperm production, which occurs within the testes. The scrotum helps to regulate the temperature of the testes and maintains it around 35 degrees Celsius (95 degrees Fahrenheit). Temperature control is accomplished by the smooth muscles of the scrotum moving the testes either closer to or further away from the abdomen, dependent upon the ambient temperature. This regulatory action is accomplished by the cremaster muscle in the abdomen and the dartos fascia (muscular tissue under the skin) within the scrotum.
The dartos muscle makes up the subcutaneous muscle layer of the scrotum (Figure 27.12). It continues internally to make up the scrotal septum, a wall that divides the scrotum into two compartments, each housing one testis. Descending from the internal oblique muscle of the abdominal wall are the two cremaster muscles, which cover each testis like a muscular net. By contracting simultaneously, the dartos and cremaster muscles can elevate the testes in cold weather (or water), moving the testes closer to the body and decreasing the surface area of the scrotum to retain heat. Alternatively, as the environmental temperature increases, the scrotum relaxes, moving the testes farther from the body core and increasing scrotal surface area, which promotes heat loss. Externally, the scrotum has a raised medial thickening on the surface called the raphe (Figure 27.12).
The testes are the male gonads —that is, the male reproductive organs. They produce both sperm and androgens, such as testosterone, and are active throughout the reproductive lifespan of the male.
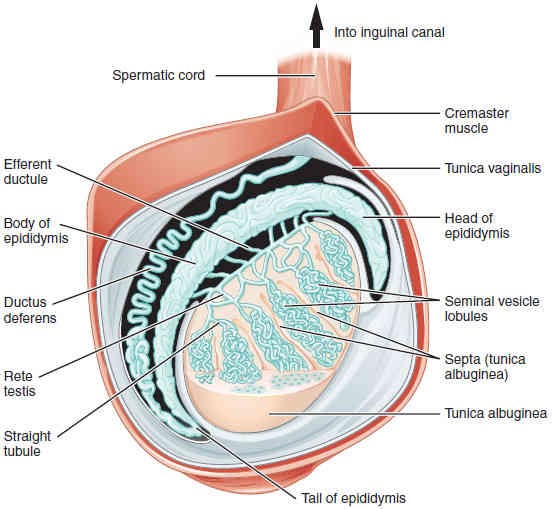
Paired ovals, the testes are each approximately 4 to 5 cm in length and are housed within the scrotum (Figures 27.11 and 27.12). They are surrounded by two distinct layers of protective connective tissue (Figure 27.13). The outer tunica vaginalis is a double-layered serous membrane. Beneath the tunica vaginalis is the tunica albuginea, a tough, white, dense connective tissue layer covering the testis itself. Not only does the tunica albuginea cover the outside of the testis, it also invaginates to form septa that divide the testis into 300 to 400 structures called seminal vesicle lobules (or just lobules). Within each lobule, sperm develop in tube-like structures known as the seminiferous tubules.
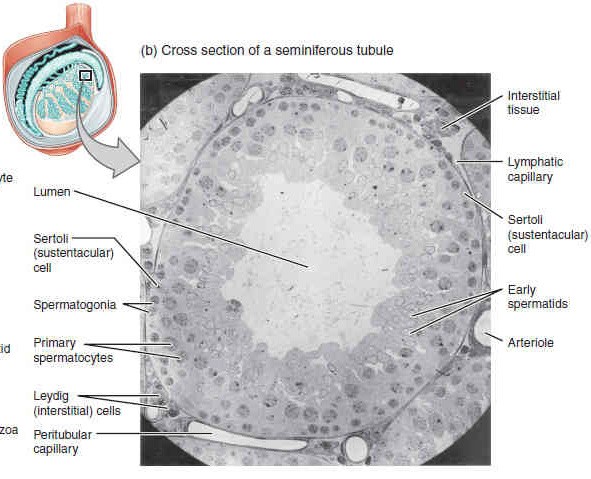
Inside the seminiferous tubules are six different cell types. These include supporting cells called sustentacular (Sertoli) cells, hormone producing interstitial (Leydig) cells, as well as five types of developing sperm cells called germ cells. Germ cell development progresses from the basement membrane—at the perimeter of the tubule—toward the lumen. Let’s look more closely at these cell types.
The least mature germ cells, the spermatogonia (singular; spermatogonium), line the basement membrane just inside the tubule. Spermatogonia are the stem cells of the testis, meaning that they are still able to differentiate into a variety of different cell types throughout adulthood. Spermatogonia initially divide to produce primary and then secondary spermatocytes, then spermatids, which will finally produce mature sperm. The process that begins with spermatogonia and concludes with the production of sperm is called spermatogenesis, which will be discussed next.
Spermiogenesis and the Structure of a Mature Sperm
Sperm are smaller than most cells in the body; in fact, the volume of a sperm cell is 85,000 times less than that of the female gamete. Approximately 100 to 300 million sperm are produced each day, whereas women typically ovulate only one oocyte per month. As is true for most cells in the body, the structure of sperm cells speaks to their function. Sperm have a distinctive head, mid-piece, and tail region (Figure 27.15).
The head of the sperm contains the extremely compact haploid nucleus with very little cytoplasm. These qualities contribute to the overall small size of the sperm (the head is only 5 μm long). A structure called the acrosome covers most of the head of the sperm cell as a “cap” that is filled with lysosomal enzymes important for preparing sperm to participate in fertilization. Tightly packed mitochondria fill the mid-piece of the sperm. ATP produced by these mitochondria will power the flagellum, which extends from the neck and the mid-piece through the tail of the sperm, enabling it to move the entire sperm cell.
Epididymis
To fertilize an egg, sperm must be moved from the seminiferous tubules in the testes, through the epididymis, and—later during ejaculation—along the length of the penis and out into the female reproductive tract.
From the lumen of the seminiferous tubules, immotile sperm are surrounded by testicular fluid and moved to the epididymis (plural; epididymides), a coiled tube attached to the testis where newly formed sperm continue to mature (Figure 4). Though the epididymis does not take up much room in its tightly coiled state, it would be approximately 6 m (20 feet) long if straightened. It takes an average of 12 days for sperm to move through the coils of the epididymis, with the shortest recorded transit time in humans being one day. Sperm enter the head of the epididymis and are moved along predominantly by the contraction of smooth muscles lining the epididymal tubes. As they are moved along the length of the epididymis, a region known as the body, the sperm further mature and acquire the ability to move under their own power. Once inside the female reproductive tract, they will use this ability to move independently toward the unfertilized egg. The more mature sperm are then stored in the tail of the epididymis (the final section) until ejaculation occurs.
Spermatic cord
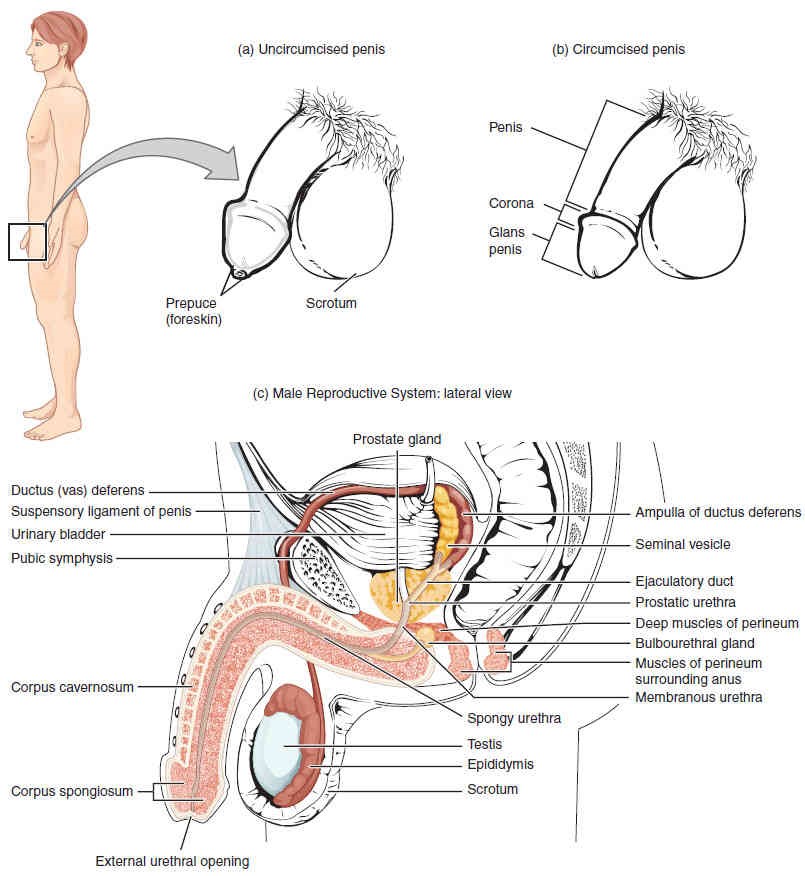
During ejaculation, sperm exit the tail of the epididymis and are pushed by smooth muscle contraction to the ductus deferens (also called the vas deferens). The vas deferens is a thick, muscular tube that is bundled together inside the scrotum with connective tissue, blood vessels, and nerves, forming a structure known as the spermatic cord (see Figure 27.11 and Figure 27.12). Since the ductus deferens is physically accessible within the scrotum, surgical sterilization to interrupt sperm delivery can be performed by cutting and sealing a small section of the ductus (vas) deferens. This procedure is called a vasectomy, and it is an effective form of male birth control.
As sperm pass through the ampulla (enlarged region) of the ductus deferens at ejaculation, they mix with fluid from the associated seminal vesicles (Figure 27.11 and Figure 27.16). The paired seminal vesicles are glands that contribute approximately 60% of the semen volume. Seminal vesicle fluid contains large amounts of fructose, which is used by the sperm mitochondria to generate ATP to allow movement through the female reproductive tract. The fluid, now containing both sperm and seminal vesicle secretions, next moves into the associated ejaculatory duct, a short structure formed from the ampulla of the ductus deferens and the duct of the seminal vesicle. The paired ejaculatory ducts transport the seminal fluid into the next structure, the prostate gland.
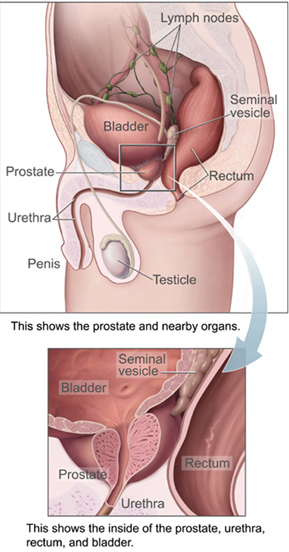
Prostate Gland
As shown in Figure 27.16, the centrally located prostate gland sits anterior to the rectum at the base of the bladder surrounding the prostatic urethra (the portion of the urethra that runs within the prostate). About the size of a walnut, the prostate is formed of both muscular and glandular tissues. It excretes an alkaline, milky fluid into the passing seminal fluid—now called semen.
The External Genitalia
The penis is the male organ of copulation (sexual intercourse). It is flaccid for non-sexual actions, such as urination, and turgid and rod-like with sexual arousal. When erect, the stiffness of the organ allows it to penetrate into the vagina and deposit semen into the female reproductive tract.
The shaft of the penis surrounds the urethra (Figure 27.17). Internally, the shaft is composed of three column-like chambers of erectile tissue that span the length of the shaft. Each of the two larger lateral chambers is the corpus cavernosum (plural; corpora cavernosa). Together, these make up the bulk of the penis. The corpus spongiosum, which can be felt as a raised ridge on the erect penis, is a smaller chamber that surrounds the spongy, or penile, urethra.
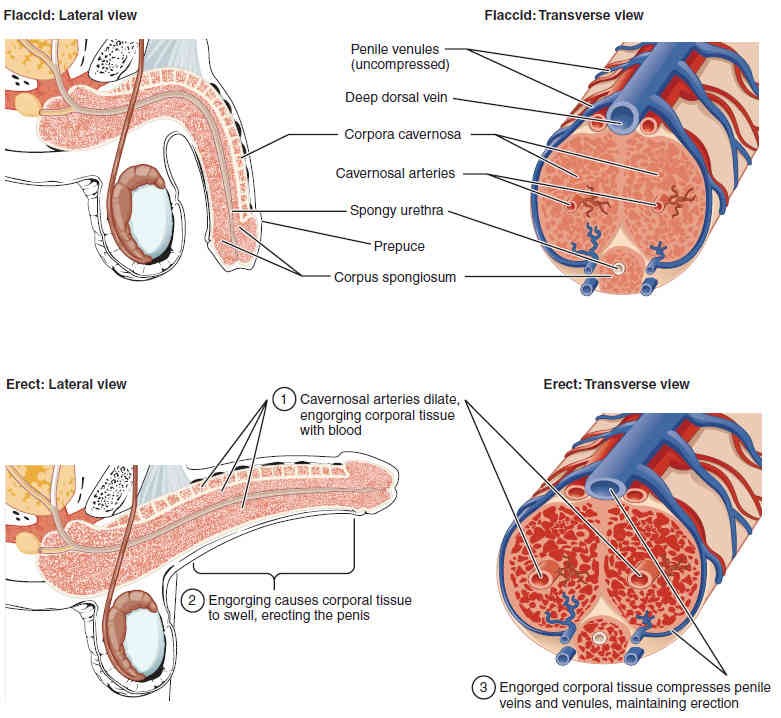
The end of the penis, called the glans penis, has a high concentration of nerve endings, resulting in very sensitive skin that influences the likelihood of ejaculation (Figure 27.11). The skin from the shaft extends down over the glans and forms a collar called the prepuce or foreskin (Figure 27.11 and Figure 27.17). The foreskin also contains a dense concentration of nerve endings, and both lubricate and protect the sensitive skin of the glans penis. A surgical procedure called circumcision, often performed for religious or social reasons, removes the prepuce, typically within days of birth.
Both sexual arousal and REM sleep (during which dreaming occurs) can induce an erection. Penile erections are the result of engorgement of the tissues because more arterial blood flows into the penile tissues than is leaving through the veins. To initiate this process during sexual arousal, nitric oxide (NO) is released from nerve endings near these blood vessels within the corpora cavernosa and spongiosum. Release of the NO activates a pathway that results in relaxation of the smooth muscles that surround the penile arteries, causing them to dilate. This dilation increases the amount of blood that can enter the penis and induces the endothelial cells in the penile arterial walls to also secrete NO and perpetuate the vasodilation. This rapid increase in blood volume fills the erectile chambers, and the increased pressure of the filled chambers compresses the thin-walled penile venules, preventing venous drainage of the penis. The result of this increased blood flow to the penis and reduced blood return from the penis is erection (Figure 27.17).
Hormones of the Male Reproductive System
Testosterone, an androgen, is a steroid hormone produced by Leydig cells. The alternate term for Leydig cells, interstitial cells, reflects their location between the seminiferous tubules in the testes. In male embryos, testosterone is secreted by Leydig cells by the seventh week of development, with peak concentrations reached in the second trimester. This early release of testosterone results in the anatomical differentiation of the male sexual organs. In childhood, testosterone concentrations are low, though they increase during puberty, activating characteristic physical changes and initiating spermatogenesis.
The continued presence of testosterone is necessary to keep the male reproductive system working properly, and Leydig cells produce approximately 6-7 mg of testosterone per day. Maintaining these normal concentrations of testosterone promotes spermatogenesis, whereas low levels of testosterone can lead to infertility. The regulation of testosterone concentrations throughout the body is critical for male reproductive function, requiring an intricate interplay between the endocrine system and the reproductive system. The relationship between these two systems is shown in Figure 27.18.
Together, the hypothalamus and pituitary gland regulate the production of testosterone and the cells that assist in spermatogenesis. Initially, gonadotropin-releasing hormone (GnRH) from the hypothalamus activates the anterior pituitary to produce luteinizing hormone (LH) and follicle stimulating hormone (FSH), which in turn stimulate Leydig cells and Sertoli cells, respectively. The system also establishes a negative feedback loop because the end products of the pathway, testosterone and inhibin, interact with the activity of GnRH to inhibit their own production (Figure 27.18, steps 2 and 3).
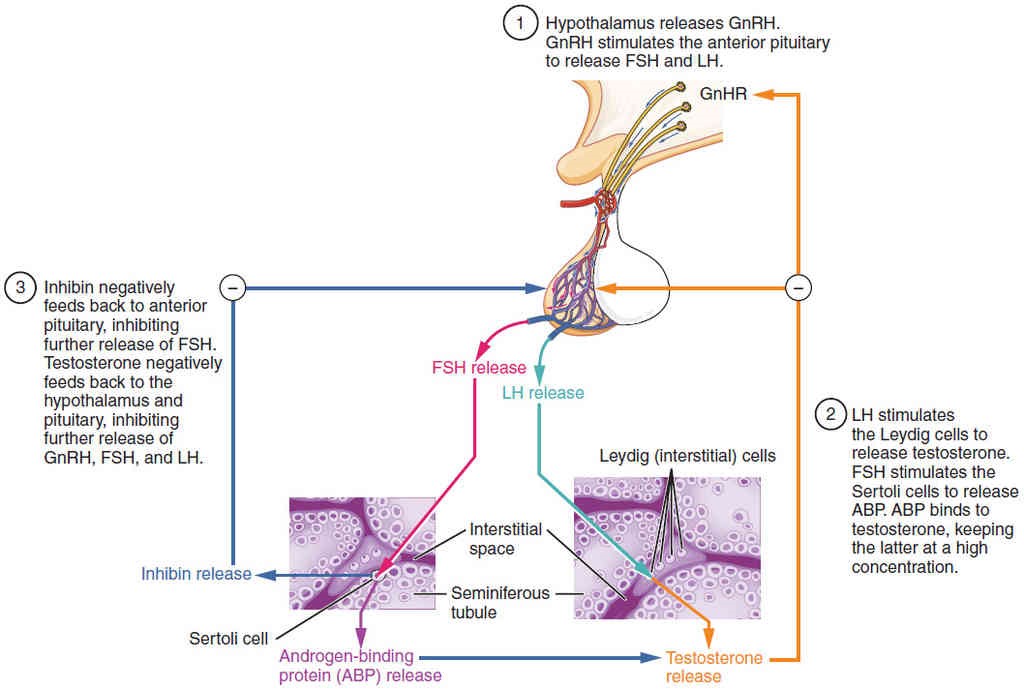
The regulation of Leydig cell production of testosterone begins outside of the testes. The hypothalamus and the pituitary gland in the brain integrate external and internal signals to control testosterone synthesis and secretion. Pulsatile release of GnRH from the hypothalamus stimulates the endocrine release of hormones from the pituitary gland. Binding of GnRH to its receptors on the anterior pituitary gland stimulates release of the two gonadotropins: LH and FSH. These two hormones are critical for reproductive function in both men and women. In men, FSH binds predominantly to the Sertoli cells within the seminiferous tubules to promote spermatogenesis. FSH also stimulates the Sertoli cells to produce hormones called inhibins, which function to inhibit FSH release from the pituitary, thus reducing testosterone secretion. In men, LH binds to receptors on Leydig cells in the testes and upregulates the production of testosterone. As previously noted, a negative feedback loop predominantly controls the synthesis and secretion of both of these hormones and testosterone.
In addition to intra-testicular secretion, testosterone is also released into the systemic circulation and plays an important role in muscle development, bone growth, the development of secondary sex characteristics, and maintaining libido (sex drive) in both males and females. In females, the ovaries secrete small amounts of testosterone, although most is converted to estradiol. A small amount of testosterone is also secreted by the adrenal glands in both sexes.
Pre-Laboratory Questions
1.What are the female gonads called?
oocytes
ova
oviducts
ovaries
2.Where does fertilization of the egg by the sperm typically occur?
vagina
uterus
uterine tube
ovary
3.The vulva includes the ________.
lactiferous duct, rugae, and hymen
lactiferous duct, endometrium, and bulbourethral glands
mons pubis, endometrium, and hymen
mons pubis, labia majora, and Bartholin’s glands
4.From what structure does the corpus luteum originate?
uterine corpus
dominant follicle
fallopian tube
corpus albicans
5.What are male gametes called?
ova
sperm
testes
testosterone
6.Which hypothalamic hormone contributes to the regulation of the male reproductive system?
luteinizing hormone
gonadotropin-releasing hormone
follicle-stimulating hormone
androgens
7.Spermatogenesis takes place in the ________.
prostate gland
glans penis
seminiferous tubules
ejaculatory duct
8.What is the function of the epididymis?
sperm maturation and storage
produces the bulk of seminal fluid
provides nitric oxide needed for erections
spermatogenesis
- Exercise 1 Overview of the female reproductive system
- Exercise 2 Microanatomy of the ovaries
- Exercise 3 Microanatomy of the uterus
- Exercise 4 Anatomy of the breast
- Exercise 5 Overview of the male reproductive system
- Exercise 6 Gross anatomy of the testes
- Exercise 7 Microanatomy of the testes
- Exercise 8 Histology of sperm
- Exercise 9 External genitalia
Exercise 1 Overview of the female reproductive system
Required Materials
- Torso models
- Female Reproductive System Poster
- Female Pelvis Models
- Human Uterus and Ovary Pathology Model
- Post-it notes
- Labeling tape
Procedure
- Look at the charts and models of the female reproductive system for a general orientation. Locate the following structures. Use the post-it notes or labeling tape to label each structure on the models. Take pictures and insert these below. Alternatively, you can sketch and label.
-
- Ovary
- Uterine (fallopian) tube
- Uterus
- Vaginal canal
- Clitoris
- Labia minora (singular, labium minus)
- Labia majora (singular, labium majus)

Exercise 2 Microanatomy of the ovaries
Required Materials
- Compound microscope
- Microscope lens paper
- Microscope lens cleaner
- Microscope immersion oil
- Slide of Human Ovary
Procedure
1.Obtain a prepared slide or a histological section of the ovary. If using a microscope, observe the sample on low power.
2.Using the slide or provided image, locate the medulla, the highly vascularized tissue in the middle of each ovary. Once identified, look for circular structures within this region. These circles are ovarian follicles. Locate the primordial follicles in your preparation. These follicles contain primary oocytes, while more mature follicles will have secondary oocytes. Sketch and label these structures below:

3.Now observe your slide under high magnification. Using your prepared sample or image, locate the primary, secondary, and tertiary follicles. Some follicles may contain oocytes. Primary follicles will have a single layer of follicular cells surrounding an oocyte; secondary follicles will have multiple layers of follicular cells surrounding an oocyte; tertiary follicles contain significant amounts of fluid in the region known as the antrum. Using your sample and Figure 27.5, try to identify a mature ovarian (Graafian) follicle; these will be the largest follicles present. Sketch and label your observations below:
Exercise 3 Microanatomy of the uterus
Required Materials
- Compound microscope
- Microscope lens paper
- Microscope lens cleaner
- Microscope immersion oil
- Slide of Human Uterus
Procedure
1.Obtain a prepared slide or a histological section of the uterus. If using a microscope, observe the sample on low power. Using the slide or provided image, locate the three layers of the uterus. Sketch and label these in the space below:
2.Now, examine the endometrium under higher power magnification. You should be able to identify two layers, including the functional layer and the basal layer. The functional layer will be the more superficial layer that is shed during menstruation, while the basal layer is deeper and will be retained. Using the same preparation or image, locate the myometrium. This layer will sit just deep to the endometrial tissue and it can be distinguished by the presence of smooth muscle. Draw an example of what you see below and label your drawing:
Exercise 4 Anatomy of the breast
Required Materials
- Female Reproductive System Poster
- Breast Cross Section Model (pathologies)
- Torso model
- Post-it notes
- Labeling tape
Procedure
- Observe the external anatomy of the breast on the provided models. The major external features of the breast include the pigmented areola, the protruding nipple, the body of the breast, and the axillary tail. Identify each of these structures in the provided materials.
- Use the provided models and charts to locate the internal structures of the breast. Much of the breast is composed of adipose tissue and embedded mammary glands. These glands are responsible for producing milk in lactating females. Identify the mammary gland and the following associated structures on the figure. Each gland consists of clusters of 15-20 lobes. Each lobe contains groups of milk-secreting cells in clusters called alveoli. These clusters can change in size depending on the amount of milk in the alveolar lumen. In nursing females, the mammary glands increase in size and lead to lactiferous ducts, which collect and direct milk to the lactiferous sinuses. Together, the ducts and sinuses collect and direct milk to exit the breast through the nipple.
- Using post-it notes or labeling tape, label these structures on the models. Take pictures and insert these in the space below. Alternatively you can sketch and label:
Exercise 5 Overview of the male reproductive system
Required Materials
- Male Reproductive System Poster
- Male Pelvis Models
- Male Pelvis with Testicular Pathology Model
- Torso models
- Post-it notes
- Labeling tape
Procedure
- Look at the charts and models of the male reproductive system for a general orientation and locate the following structures. Use the post-it notes or labeling tape to label each structure on the models. Take pictures and insert these below. Alternatively, you can sketch and label.
- Testis
- Epididymis
- Scrotal sac (scrotum)
- Ductus (vas) deferens
- Seminal vesicle
- Prostate gland
- Bulbourethral gland
- Penis
- Urethra
Exercise 6 Gross anatomy of the testes
Required Materials
- Male Reproductive System Poster
- Male Pelvis Models
- Male Pelvis with Testicular Pathology Model
- Torso models
- Post-it notes
- Labeling tape
Procedure
- Examine charts and a model of the testes. The testes are paired organs, sitting outside of the body.
- Using the models identify the tunica albuginea, a tough connective sheath that surrounds the testes. Locate the invaginations of the membrane, where it invaginates to form many lobules within each testis. Located superficial to the tunica, is the scrotal sac (scrotum), this structure keeps the testes on the exterior of the body, where temperature tends to be cooler and more supportive to spermatozoa (sperm) production. Locate the dartos muscle, a component of the scrotal sac. When the testes are cold, the muscle contracts, tightening the sac and bringing the testes closer to the body. The opposite actions occur when the environment is warm.
- Use the post-it notes or labeling tape to label each structure on the models. Take pictures and insert these below. Alternatively, you can sketch and label.
Exercise 7 Microanatomy of the testes
Required Materials
- Compound microscope
- Microscope lens paper
- Microscope lens cleaner
- Microscope immersion oil
- Slide of Human Testis
- Model of Meiosis
Procedure
- Obtain a prepared slide or a histological section of the testes. Identify the seminiferous tubules . Multiple tubules may be identifiable in the preparation. It is within these structures where sperm are produced. Look for the triangular clusters of cells in between each tubule. These are the interstitial (Leydig) cells. They will produce the male sex hormone, testosterone. Sketch and label these tubules and interstitial structures in the space below:
2. Examine the seminiferous tubules under high magnification. You should be able to see an outer row of cells, known as the spermatogonia. These cells will divide by the process of mitosis, giving rise to primary spermatocytes. The primary spermatocytes will then undergo meiosis, or reduction division, to eventually produce spermatozoa. To do so, primary spermatocytes will initially divide to form secondary spermatocytes, which are found closer to the lumen of the seminiferous tubules. These cells will then become spermatids. Ultimately, the spermatids will lose their remaining cytoplasm and mature into functional spermatozoa. Locate the primary and secondary spermatocytes, spermatids, and spermatozoa. You may be able to see sustentacular (Sertoli) cells, which help nourish, support, and move the sperm during development.
3. Draw an example of what you see at high magnification of what is listed in step 2 in the space provided below:
Exercise 8 Histology of sperm
Required Materials
- Compound microscope
- Microscope lens paper
- Microscope lens cleaner
- Microscope immersion oil
- Slide of Human Sperm
Procedure
- Examine a prepared slide of sperm.
- Identify the different components of the sperm cell. Each sperm consists of a head, midpiece, and tail. Sketch and label these structures below:
Exercise 9 External genitalia
Required Materials
- Male Reproductive System Poster
- Male Pelvis Models
- Male Pelvis with Testicular Pathology Model
- Torso Models.
- Post-it notes
- Labeling tape
Procedure
- In addition to the testes, which are housed within the scrotum, the penis is considered to contribute to the external genitalia of a male’s reproductive system. Use the provided models to locate and observe the external anatomy of the penis. The penis consists of the root, bulb, an elongated shaft, and a distally expanded region known as that glans penis. The glans is covered with the prepuce, or foreskin. Identify and label all of these structures using post-it notes or labeling tape. Take pictures and insert below. Alternatively, you can sketch and label.
- Now, examine a model or chart of a cross section of the penis. Notice that the penis contains three distinct cylinders of erectile tissue that are anchored to the body proximally. Identify the corpus spongiosum, the cylinder of erectile tissue that contains the spongy (penile) urethra. Located dorsal to the spongiosum, two cylinders of corpus cavernosa are located. Follow the corpus spongiosum as it extends distally. At the most distal region of the penis, this tissue expands to form the glans penis. Note the dorsal arteries and deep (cavernosal) arteries. Together, these vessels take blood to the penis. When these vessels dilate, the erectile tissue of the penis engorge with blood, making the penis erect. Also locate the dorsal vein and venules of the penis. These vessels will remove blood from the penis, except for when the penis is erect. At this point, these vessels are compressed, preventing venous drainage of the penis.
- Sketch and label all of the structure listed in step 2 in the space below:
Post-laboratory Questions
- The __________________ is the inner epithelial lining of the uterus.
- A follicle is comprised of _________________ cells, _____________ cells, and the _____________.
- The finger-like structures on the fallopian tube that help sweep the ovum into the ampulla are called __________________.
- The layer of cells surrounding the ovulated ovum are called the _________________________.
- The _______________________ holds the uterus and ovaries in place within the body.
- The ____________________________ is the layer of the endometrium that is shed every month.
- The ___________ is a tube that allows sperm cells to travel from the testes to the urethra.
- Which cells are responsible for the production of testosterone in response to LH from the anterior pituitary?
- The _____________, _____________, and _____________, provide important fluids to the sperm before ejaculation.
- True or False: warmer temperatures (above body temp) are essential for spermatogenesis.
